The concept of inheritance of traits is widely studied in genetics. It is they who explain the similarity between offspring and parents. It is curious that some manifestations of traits are inherited together. This phenomenon, first described in detail by the scientist T. Morgan, came to be called “linked inheritance.” Let's talk about it in more detail.
As you know, each organism has a certain number of genes. At the same time, chromosomes are also a strictly limited number. For comparison: a healthy human body has 46 chromosomes. There are thousands of times more genes in it. Judge for yourself: each gene is responsible for one or another trait that manifests itself in a person’s appearance. Naturally, there are a lot of them. Therefore, they began to talk about the fact that several genes are localized on one chromosome. These genes are called a linkage group and determine linked inheritance. A similar theory has been floating around in the scientific community for quite a long time, but only T. Morgan gave it a definition.
Unlike the inheritance of genes that are localized in different pairs of identical chromosomes, linked inheritance causes a diheterozygous individual to form only two types of gametes, repeating the combination of parental genes.
Along with this, gametes arise, the combination of genes in which differs from the chromosomal set of the parents. This result is a consequence of crossing over, a process whose importance in genetics is difficult to overestimate, since it allows the offspring to receive different traits from both parents.
In nature, there are three types of gene inheritance. In order to determine which type is inherent in a given pair of them, they are used. The result will necessarily result in one of the three options given below:
1. Independent inheritance. In such a case, hybrids differ from each other and from their parents in appearance, in other words, as a result we have 4 variants of phenotypes.
2. Complete linkage of genes. First generation hybrids, resulting from crossing parental individuals, completely repeat the phenotype of the parents and are indistinguishable from each other.
3. Incomplete linkage of genes. Just as in the first case, when crossed, 4 classes of different phenotypes are obtained. In this case, however, new genotypes are formed that are completely different from the parent stock. It is in this case that crossing over, mentioned above, interferes with the process of gamete formation.
It has also been established that the smaller the distance between inherited genes on the parent chromosome, the higher the likelihood of their complete linked inheritance. Accordingly, the farther they are located from each other, the less often crossover occurs during meiosis. The distance between genes is the factor that primarily determines the probability of linked inheritance.
Separately, it is necessary to consider linked inheritance associated with gender. Its essence is the same as with the option discussed above, however, the inherited genes in this case are located on the sex chromosomes. Therefore, we can talk about this type of inheritance only in the case of mammals (including humans), some reptiles and insects.
Taking into account the fact that XY is a set of chromosomes corresponding to the male sex, and XX to the female sex, we note that all the main characteristics responsible for the viability of the organism are located in the chromosome present in the genotype of each organism. Of course, we are talking about the X chromosome. In females, both recessive and chromosomal ones may be present. Males can inherit only one of the variants - that is, either the gene manifests itself in the phenotype or not.
Sex-linked inheritance is often heard in the context of diseases that are specific to men, while women are only their carriers:
In 1906, W. Batson and R. Punnett, crossing sweet pea plants and analyzing the inheritance of pollen shape and flower color, discovered that these characteristics do not give independent distribution in the offspring; hybrids always repeated the characteristics of the parent forms. It became clear that not all traits are characterized by independent distribution in the offspring and free combination.
Each organism has a huge number of characteristics, but the number of chromosomes is small. Consequently, each chromosome carries not one gene, but a whole group of genes responsible for the development of different traits. He studied the inheritance of traits whose genes are localized on one chromosome. T. Morgan. If Mendel conducted his experiments on peas, then for Morgan the main object was the fruit fly Drosophila.
Drosophila produces numerous offspring every two weeks at a temperature of 25 °C. The male and female are clearly distinguishable in appearance - the male has a smaller and darker abdomen. They have only 8 chromosomes in a diploid set and reproduce quite easily in test tubes on an inexpensive nutrient medium.
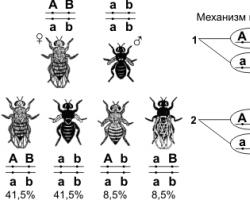
By crossing a Drosophila fly with a gray body and normal wings with a fly having a dark body color and rudimentary wings, in the first generation Morgan obtained hybrids with a gray body and normal wings (the gene that determines the gray color of the abdomen dominates the dark color, and the gene that determines development of normal wings, - above the gene of underdeveloped wings). When carrying out an analytical crossing of an F 1 female with a male who had recessive traits, it was theoretically expected to obtain offspring with combinations of these traits in a ratio of 1:1:1:1. However, in the offspring, individuals with characteristics of the parental forms clearly predominated (41.5% - gray long-winged and 41.5% - black with rudimentary wings), and only a small part of the flies had a combination of characters different from those of the parents (8.5% - black long-winged and 8.5% - gray with rudimentary wings). Such results could only be obtained if the genes responsible for body color and wing shape are located on the same chromosome.

1 - non-crossover gametes; 2 - crossover gametes.
If the genes for body color and wing shape are localized on one chromosome, then this crossing should have resulted in two groups of individuals repeating the characteristics of the parental forms, since the maternal organism should form gametes of only two types - AB and ab, and the paternal one - one type - ab . Consequently, two groups of individuals with the genotype AABB and aabb should be formed in the offspring. However, individuals appear in the offspring (albeit in small numbers) with recombined traits, that is, having genotypes Aabb and aaBb. In order to explain this, it is necessary to recall the mechanism of formation of germ cells - meiosis. In the prophase of the first meiotic division, homologous chromosomes are conjugated, and at this moment an exchange of regions can occur between them. As a result of crossing over, in some cells, sections of chromosomes are exchanged between genes A and B, gametes Ab and aB appear, and, as a result, four groups of phenotypes are formed in the offspring, as with the free combination of genes. But, since crossing over occurs during the formation of a small part of gametes, the numerical ratio of phenotypes does not correspond to the ratio 1:1:1:1.
Clutch group- genes localized on the same chromosome and inherited together. The number of linkage groups corresponds to the haploid set of chromosomes.
Chained inheritance- inheritance of traits whose genes are localized on the same chromosome. The strength of linkage between genes depends on the distance between them: the further the genes are located from each other, the higher the frequency of crossing over and vice versa. Full grip- a type of linked inheritance in which the genes of the analyzed traits are located so close to each other that crossing over between them becomes impossible. Incomplete clutch- a type of linked inheritance in which the genes of the analyzed traits are located at a certain distance from each other, which makes crossing over between them possible.
Independent inheritance- inheritance of traits whose genes are localized in different pairs of homologous chromosomes.
Non-crossover gametes- gametes during the formation of which crossing over did not occur.
Non-recombinants- hybrid individuals that have the same combination of characteristics as their parents.
Recombinants- hybrid individuals that have a different combination of characteristics than their parents.
The distance between genes is measured in Morganids— conventional units corresponding to the percentage of crossover gametes or the percentage of recombinants. For example, the distance between the genes for gray body color and long wings (also black body color and rudimentary wings) in Drosophila is 17%, or 17 morganids.
In diheterozygotes, dominant genes can be located either on one chromosome ( cis phase), or in different ( trans phase).

1 - Cis-phase mechanism (non-crossover gametes); 2 - trans-phase mechanism (non-crossover gametes).
The result of T. Morgan's research was the creation of chromosomal theory of heredity:
Go to lectures No. 17“Basic concepts of genetics. Mendel's laws"
Linked inheritance is the phenomenon of correlated inheritance of certain states of genes located on the same chromosome.
There is no complete correlation due to meiotic crossing over, since linked genes can disperse into different gametes. Crossing over is observed in the form of unlinking in the offspring of those gene alleles and, accordingly, states of traits that were linked in the parents.
Observations made by Thomas Morgan showed that the probability of crossing over between different pairs of genes varied, and the idea came to create gene maps based on the frequencies of crossing over between different genes. The first gene map was constructed by Morgan's student, Alfred Sturtevant, in 1913 using material from Drosophila melanogaster.
The distance between genes located on the same chromosome is determined by the percentage of crossing over between them and is directly proportional to it. The unit of distance is taken to be 1% crossing over (1 morganid or 1 centimorganid). The farther genes are from each other on a chromosome, the more often crossing over will occur between them. The maximum distance between genes located on the same chromosome can be 49 centimorganides.
Linked traits
Linked traits are traits that are controlled by genes located on the same chromosome. Naturally, they are transmitted together in cases of complete coupling.
Morgan's Law
Linked genes localized on the same chromosome are inherited together and do not exhibit independent distribution
Crossing over
However, homologous chromosomes can cross over (crossing over or decussation) and exchange homologous regions. In this case, the genes of one chromosome are transferred to another, homologous to it. The closer the genes are located to each other on a chromosome, the stronger the linkage between them and the less often their divergence occurs during crossing over, and, conversely, the farther the genes are from each other, the weaker the linkage between them and the more often its disruption is possible.
The number of different types of gametes will depend on the frequency of crossing over or the distance between the genes analyzed. The distance between genes is calculated in morganids: one unit of distance between genes located on the same chromosome corresponds to 1% crossing over. This relationship between distances and crossing-over frequency can be traced only up to 50 morganids.
The chromosomal theory of heredity is a theory according to which chromosomes contained in the cell nucleus are carriers of genes and represent the material basis of heredity, that is, the continuity of the properties of organisms in a number of generations is determined by the continuity of their chromosomes. The chromosomal theory of heredity arose at the beginning of the 20th century. based on cell theory and was used to study the hereditary properties of organisms through hybridological analysis.
The founder of the chromosome theory, Thomas Gent Morgan, American geneticist, Nobel laureate. Morgan and his students found that:
– each gene has a specific locus (location) on the chromosome;
– genes on a chromosome are located in a certain sequence;
– the most closely located genes on one chromosome are linked, therefore they are inherited predominantly together;
– groups of genes located on the same chromosome form linkage groups;
– the number of linkage groups is equal to the haploid set of chromosomes in homogametic individuals and n+1 in heterogametic individuals;
– an exchange of sections (crossing over) can occur between homologous chromosomes; as a result of crossing over, gametes arise whose chromosomes contain new combinations of genes;
– the frequency (in%) of crossing over between non-allelic genes is proportional to the distance between them;
– the set of chromosomes in cells of a given type (karyotype) is a characteristic feature of the species;
– the frequency of crossing over between homologous chromosomes depends on the distance between genes localized on the same chromosome. The greater this distance, the higher the crossing over frequency. The unit of distance between genes is taken to be 1 morganid (1% crossing over) or the percentage of occurrence of crossover individuals. If this value is 10 morganids, it can be stated that the frequency of chromosome crossings at the locations of these genes is 10% and that new genetic combinations will be identified in 10% of the offspring.
To clarify the nature of the location of genes on chromosomes and determine the frequency of crossing over between them, genetic maps are constructed. The map reflects the order of genes on a chromosome and the distance between genes on the same chromosome. These conclusions of Morgan and his colleagues were called the chromosomal theory of heredity. The most important consequences of this theory are modern ideas about the gene as a functional unit of heredity, its divisibility and ability to interact with other genes.
The formation of the chromosome theory was facilitated by data obtained from the study of the genetics of sex, when differences in the set of chromosomes were established in organisms of different sexes.
Mendel's third law of independent inheritance is violated if the genes are on the same chromosome. For the first time, the phenomenon of linked genes, i.e. located on the same chromosome, was observed by the geneticist Thomas Morgan. Subsequently, the simultaneous inheritance of two characteristics was called Morgan's law.
To talk briefly and clearly about Thomas Morgan's law, you should first remember what a chromosome is.
A chromosome is a structure located in the cell nucleus that carries hereditary information. It consists of a long chain of DNA, which in turn consists of genes - units of hereditary information. Each gene is responsible for a specific trait. The set of chromosomes is called a karyotype.

Rice. 1. Chromosome.
Mendel looked at traits found on different chromosomes. When crossing, different combinations of genes are formed that form the genotype of the individual.
Unlike Mendel's law, Morgan's law applies to genes located on the same chromosome.
The wording of the law is as follows: genes located close to each other on the same chromosome form a group and are inherited in a linked manner. The number of linked groups corresponds to the haploid set - half of the complete set of chromosomes. Humans have 46 chromosomes, i.e. 23 pairs, respectively 23 clutch groups.
TOP 3 articleswho are reading along with this

Rice. 2. Morgan's Law.
The frequency of inheritance depends on the distance between genes. The closer the genes that form groups are, the more often linked traits are inherited, i.e. When placed close together, the adhesion force is stronger.
Examples of chained inheritance:
If the genes are not linked, then four types of gametes AaBb are formed - AB, aB, Ab, ab. When crossing hybrids, the phenotypic ratio will be 9:3:3:1 (cleavage will occur). With linked inheritance, two types of gametes are formed - AB and ab. In this case, the F2 generation will produce offspring with a 3:1 phenotype.
T. Morgan's law of linked inheritance may be violated. Gene sections are exchanged between homologous chromosomes, and new gene combinations are formed. This phenomenon is called crossing over. Disruption of connections occurs in meiosis during conjugation (prophase I - approaching and joining). Homologous chromosomes exchange sections, breaking the linked bonds. In this case, Mendel's third law is fully observed.

Rice. 3. Crossing over.
Morgan's work was as follows:
The possibility of crossing over increases if linked genes are located at a considerable distance from each other. The lower the percentage of crossing over, the greater the likelihood of linked inheritance.
Chained inheritance. Independent distribution of genes (Mendel's second law) is based on the fact that genes belonging to different alleles are located in different pairs of homologous chromosomes. The question naturally arises: how will the distribution of different (non-allelic) genes occur in a number of generations if they lie in the same pair of chromosomes? This phenomenon must take place, because the number of genes is many times greater than the number of chromosomes. Obviously, the law of independent distribution (Mendel's second law) does not apply to genes located on the same chromosome. It is limited only to those cases where the genes of different alleles are located on different chromosomes.
Pattern of inheritance when genes are found on the same chromosome, it was carefully studied by T. Morgan and his school. The main object of research was the small fruit fly Drosophila
This insect is extremely convenient for genetic work. The fly is easily bred in the laboratory, produces a new generation every 10–15 days at its optimal temperature of 25–26° C, has numerous and varied hereditary characteristics, and has a small number of chromosomes (8 in the diploid set).
Experiments have shown that genes localized on one chromosome are linked, i.e., they are inherited predominantly together, without showing independent distribution. Let's look at a specific example. If you cross a Drosophila with a gray body and normal wings with a fly that has a dark body color and rudimentary wings, then in the first generation all the flies will be gray with normal wings. This is a heterozygote for two pairs of alleles (gray body - dark body and normal wings - rudimentary wings). Let's crossbreed. Let us cross the females of these diheterozygous flies (gray body and normal wings) with males possessing recessive traits - a dark body and rudimentary wings. Based on the second, one would expect to obtain four flies in the offspring: 25% gray, with normal wings; 25% gray, with rudimentary wings; 25% dark, with normal wings; 25% dark, with rudimentary wings.
In fact, in the experiment there are significantly more flies with the original combination of characteristics (gray body - normal wings, dark body - rudimentary wings) (in this experiment, 41.5%) than flies with recombined characters (gray body - rudimentary wings and dark body - normal wings).
There will be only 8.5% of each type. This example shows that the genes that determine the characteristics of a gray body - normal wings and a dark body - rudimentary wings are inherited predominantly together, or, in other words, turn out to be linked with each other. This linkage is a consequence of the localization of genes on the same chromosome. Therefore, during meiosis, these genes do not separate, but are inherited together. The phenomenon of linkage of genes localized on the same chromosome is known as Morgan's law.
Why, after all, among the second generation hybrids do a small number of individuals appear with recombination of parental characteristics? Why is gene linkage not absolute? Research has shown that this recombination of genes is due to the fact that during the process of meiosis, during the conjugation of homologous chromosomes, they sometimes exchange their sections, or, in other words, crossover occurs between them.

It is clear that in this case, genes that were originally located on one of two homologous chromosomes will end up on different homologous chromosomes. There will be a recombination between them. The frequency of crossover is different for different genes. It depends on the distance between them. The closer the genes are located on the chromosome, the less often they are separated during crossover. This happens because chromosomes exchange different regions, and genes that are closely located are more likely to end up together. Based on this pattern, it was possible to construct genetic maps of chromosomes for well-studied organisms, on which the relative distance between genes is plotted.
The biological significance of chromosome crossing is very great. Thanks to it, new hereditary combinations of genes are created, hereditary variability increases, which supplies material for.
Genetics of sex. It is well known that in dioecious organisms (including humans) the sex ratio is usually 1:1. What reasons determine the sex of a developing organism? This question has long been of interest to humanity due to its great theoretical and practical significance. The chromosome set of males and females in most dioecious organisms is not the same. Let's get acquainted with these differences using the example of the set of chromosomes in Drosophila.
Males and females do not differ from each other in three pairs of chromosomes. But for one couple there are significant differences. The female has two identical (paired) rod-shaped chromosomes; The male has only one such chromosome, the pair of which is a special, double-armed chromosome. Those chromosomes for which there are no differences between males and females are called autosomes. The chromosomes on which males and females differ from each other are called sex chromosomes. Thus, the chromosome set of Drosophila consists of six autosomes and two sex chromosomes. The sex, rod-shaped chromosome, present in a double number in a female and single in a male, is called the X chromosome; the second, sexual (two-armed chromosome of the male, absent in the female) - the Y chromosome.
How are the considered sex differences in the chromosome sets of males and females maintained in the process? To answer this question, it is necessary to clarify the behavior of chromosomes in meiosis and during fertilization. The essence of this process is presented in the figure.

During the maturation of germ cells in a female, each egg cell, as a result of meiosis, receives a set of four chromosomes: three autosomes and one X chromosome. Males produce two types of sperm in equal quantities. Some carry three autosomes and an X chromosome, others carry three autosomes and a Y chromosome. During fertilization, two combinations are possible. An egg can be equally likely to be fertilized by a sperm with an X or Y chromosome. In the first case, a female will develop from a fertilized egg, and in the second, a male. The sex of an organism is determined at the time of fertilization and depends on the chromosome complement of the zygote.
In humans, the chromosomal mechanism for determining sex is the same as in Drosophila. The diploid number of human chromosomes is 46. This number includes 22 pairs of autosomes and 2 sex chromosomes. In women there are two X chromosomes, in men there is one X and one Y chromosome.

Accordingly, men produce sperm of two types - with X- and Y-chromosomes.
In some dioecious organisms (for example, some insects), the Y chromosome is completely absent. In these cases, the male has one less chromosome: instead of the X and Y chromosomes, he has one X chromosome. Then, during the formation of male gametes during meiosis, the X chromosome does not have a partner for conjugation and goes into one of the cells. As a result, half of all sperm have an X chromosome, while the other half lack it. When an egg is fertilized by sperm with an X chromosome, a complex with two X chromosomes is obtained, and a female develops from such an egg. If an egg is fertilized by a sperm without an X chromosome, an organism with one X chromosome (received through the egg from the female) will develop, which will be a male.
In all the examples discussed above, sperm of two categories develop: either with the X and Y chromosomes (Drosophila, humans), or half of the sperm carry the X chromosome, and the other is completely devoid of it. Eggs are all the same in terms of sex chromosomes. In all these cases we have male heterogamety (multigametic). The female sex is homogametic (equal gametic). Along with this, another type of sex determination occurs in nature, characterized by female heterogamety. Here the opposite relationships to those just discussed take place. Different sex chromosomes or only one X chromosome are characteristic of the female sex. The male sex has a pair of identical X chromosomes. Obviously, in these cases, female heterogamety will occur. After meiosis, two types of egg cells are formed, while with regard to the chromosomal complex, all sperm are the same (all carry one X chromosome). Consequently, the sex of the embryo will be determined by which egg - with the X or Y chromosome - will be fertilized.