Иннервация слезной и слюных желез
Афферентным путем для слезной железы является n. lacrimalis (ветвь n. ophthalmicus от n. trigemini), для поднижнечелюстной и подъязычной - n. lingualis (ветвь n. mandibularis от n. trigemini) и chorda tympani (ветвь n. intermedius), для околоушной - n. auriculotemporal и n. glossopharyngeus.
Эфферентная парасимпатическая иннервация слезной железы. Центр лежит в верхнем отделе продолговатого мозга и связан с ядром промежуточного нерва (nucleus salivatorius superior). Преганглионарные волокна идут в составе n. intermedius, далее n. petrosus major до ganglion pterygopalatinum. Отсюда начинаются постганглионарные волокна, которые в составе n. maxillaris и далее его ветви, n. zygoma ticus, через связи с n. lacrimalis достигают слезной железы.
Эфферентная парасимпатическая иннервация поднижнечелюстной и подъязычной желез. Преганглионарные волокна идут от nucleus salivatorius superior в составе n. intermedius, далее chorda tympani и n. lingualis до ganglion submandibulare, откуда начинаю гея поспан-глионарные волокна, достигающие желез.
Эфферентная парасимпатическая иннервация околоушной железы. Преганглионарные волокна идут от nucleus salivatorius inferior в составе n. glossopharyngeus, далее n. tympanicus, n. petrosus minor до ganglion oticum. Отсюда начинаются постганглионарные волокна, идущие к железе в составе n. auriculotemporalis. Функция: усиление секреции слезной и названных слюнных желез; расширение сосудов желез.
Эфферентная симпатическая иннервация всех названных желез. Преганглионарные волокна начинаются в боковых рогах верхних грудных сегментов спинного мозга и заканчиваются в верхнем шейном узле симпатического ствола. Постганглионарные волокна начинаются в названном узле и доходят до слезной железы в составе plexus caroticus internus, до околоушной - в составе plexus caroticus externus и до поднижнечелюстной и подъязычной желез - через plexus caroticus externus и затем через plexus facialis.
По поводу малых слюнных желез ничего нигде нет, но! они располагаются в слизистой полости рта, которая иннервируется ветямии нижнего альвеолярного нерва (n. alveolaris inferior ) (- нижнечелюстной нерв - тройничный нерв) , а так как слизистая иннервируется тройничным нервом, как и все остальные железы, то дальше информация пойдет аналогично другим структурам.
Билет 48.
1. Костно-фиброзные каналы (удерживатели сгибателей и разгибателей, каналы запястья), влагалища (синовиальные) сухожилий мышц верхней конечности. Синовиальные сумки. РАЗГИБАТЕЛИ
Подкожная жировая клетчатка задней области запястья рыхлая, развита умеренно. В ней легко скапливается отечная жидкость. Собственная фасция тыльной поверхности запястья утолщена и образует удерживателъ разгибателей, retinaculum musculorum extensoram. Под ним располагаются 6 костно-фиброзных каналов, образующихся в результате отхождения от retinaculum mm. extensoram фасциальных перегородок, прикрепляющихся к костям и связкам запястья. В каналах располагаются сухожилия мышц - разгибателей запястья и пальцев, окруженные синовиальными влагалищами.
Начиная с медиальной (локтевой) стороны, это следующие каналы: 1. Канал локтевого разгибателя запястья, m. extensor carpi ulnaris. Его синовиальное влагалище простирается от головки локтевой кости до прикрепления сухожилия к основанию V пястной кости. 2. Канал разгибателя мизинца, m. extensor digiti minimi. Синовиальное влагалище разгибателя мизинца проксимально находится на уровне дистального лучелоктевого сустава, а дистально - ниже середины V пястной кости. 3. Канал сухожилий m. extensor digitorum и m. extensor indicis, заключенных в треугольное синовиальное влагалище с основанием, обращенным в сторону пальцев 4. Канал m. extensor pollicis longus. Сухожилие этой мышцы, находящееся в собственном синовиальном влагалище, vagina tendinis m. extensoris pollicis longi, поворачивает под острым углом в латеральную сторону и пересекает спереди сухожилия лучевых разгибателей кисти, mm. extensores carpi radiales longus et brevis. 5. Костно-фиброзный канал лучевых разгибателей кисти, mm. extensores carpi longus et brevis, находится латеральнее и глубже предыдущего. Синовиальные влагалища сухожилий этих мышц могут сообщаться с полостью лучезапястного сустава. 6. Канал m. abductor pollicis longus и m. extensor pollicis brevis находится на латеральной поверхности шиловидного отростка лучевой кости.
СГИБАТЕЛИ Синовиальные влагалища на ладонной поверхности заключают: первое - сухожилия поверхностного и глубокого сгибателей пальцев, второе - длинный сгибатель I пальца. Оба синовиальных влагалища располагаются в запястном канале (canalis carpalis), который ограничен костями запястья и retinaculum flexorum. Вверху синовиальные влагалища распространяются на 1-1,5 см выше retinaculum flexorum. Внизу первое влагалище образует расширение в области сухожилий II, III, IV пальцев, заканчиваясь на середине пястных костей. Синовиальное влагалище, окружающее сухожилие сгибателя V пальца, начинается от уровня лучезапястного сустава и достигает дистальной фаланги V пальца. II, III и IV пальцы имеют самостоятельные синовиальные влагалища для сухожилий поверхностного и глубокого сгибателей пальцев. Второе синовиальное влагалище для сухожилия длинного сгибателя I пальца распространяется до дистальной фаланги.Синовиальная сумка (лат. bursa synovialis ) - небольшая уплощённой формы полость, выстланная синовиальной оболочкой, отграниченная от окружающих тканей капсулой и заполненная синовиальной жидкостью. По расположению различают подкожные, подфасциальные, подсухожильные и подмышечные синовиальные сумки.1Синовиальные сумки верхней конечности, bursae membri superiris .2Подсухожильная сумка трапециевидной мышцы, b. subtendinea m.trapezii . Локализуется между восходящей частью m. trapezius и остью лопатки. 3Акромиальная подкожная сумка, b. subcutanea acromialis 4Подакромиальная сумка, b. subacromialis . Находится под акромионом и дельтовидной мышцей на капсуле плечевого сустава. 5Поддельтовидная сумка, b. subdeltoidea . Расположена между дельтовидной мышцей и капсулой плечевого сустава. Иногда соединяется с подакромиальной сумкой6Сумка клювоплечевой мышцы, b. m.coracobrachialis . Локализуется ниже верхушки клювовидного отростка между сухожилиями подлопаточной и клювоплечевой мышц. 7Подсухожильная сумка подостной мышцы, b. subtendinea m. infraspinati . Находится между сухожилием подостной мышцы и капсулой плечевого сустава. 8Подсухожильная сумка подлопаточной мышцы, b. subtendinea m. subscapularis . Расположена между сухожилием подлопаточной мышцы и капсулой плечевого сустава. Соединяется с суставной полостью. 9Подсухожильная сумка большой круглой мышцы, b. subtendinea m. teretis majoris . Расположена между сухожилием соответствующей мышцы и плечевой костью. 10Подсухожильная сумка широчайшей мышцы спины, b. subtendinea m. latissimi dorsi . Находится между сухожилиями большой круглой мышцы и широчайшей мышцей спины11Локтевая подкожная сумка, b.subcutanea olecrani . Находится между локтевым отростком и кожей. 12Локтевая внутрисухожильная сумка, b.intratendinea olecrani . Расположена внутри сухожилия трехглавой мышцы плеча, вблизи локтевого отростка. 13Подсухожильная сумка трехглавой мышцы плеча, b. subtendinea m. tricipitis brachii . Находится между сухожилием одноименной мышцы и локтевым отростком. 14Двуглаво-лучевая сумка, b. bicipitoradialis . Локализуется между сухожилием двуглавой мышцы и бугристостью лучевой кости. 15Межкостная локтевая сумка, b.cubitalis interossea . Находится между сухожилием двуглавой мышцы и локтевой костью или косой хордой.
К большим слюнным железам (glandulae salivariae majores) относят парные околоушную, подъязычную и поднижнечелюстную железы .
Большие слюнные железы относятся к паренхиматозным органам, в составе которых выделяют:
паренхиму - специализированную (секреторную) часть железы, представленную ацинарным отделом, содержащим секреторные клетки, где происходит выработка секрета. В состав слюнных желез входят слизистые клетки, выделяющие густой слизистый секрет, и серозные клетки, выделяющие жидкую, водянистую, так называемую серозную или белковую слюну. Вырабатываемый в железах секрет по системе выводных протоков доставляется на поверхность слизистой оболочки в разных отделах полости рта.
строму - комплекс соединительнотканных структур, образующих внутренний каркас органа и способствующих формированию долек и долей; в прослойках соединительной ткани проходят сосуды и нервы, направляющиеся к ацинарным клеткам.
Околоушная железа (glandula parotidea) - наиболее крупная из слюнных желез, которая располагается книзу и кпереди от ушной раковины, у заднего края жевательной мышцы. Здесь она легко доступна для прощупывания.
Иногда может присутствовать также добавочная околоушная железа (glandula parotidea accessoria), расположенная на поверхности жевательной мышцы около протока околоушной железы. Околоушная железа - сложная многодольчатая альвеолярная железа, состоящая из серозных клеток, вырабатывающих серозную (белковую) слюну. В ней различают поверхностную часть (pars superficialis) и глубокую часть (pars profunda).
Поверхностная часть железы имеет жевательный отросток и располагается на ветви нижней челюсти и на жевательной мышце. Иногда также встречается верхний отросток, прилежащий к хрящевому отделу наружного слухового хода. Глубокая часть часто имеет глоточный и задний отростки. Она расположена в занижнечелюстной ямке (fossa retromandibularis), где прилежит к височно-нижнечелюстному суставу, сосцевидному отростку височной кости и некоторым мышцам шеи.
Околоушная железа покрыта околоушной фасцией, образующей капсулу железы. Капсула состоит из поверхностного и глубокого листков, покрывающих железу снаружи и изнутри. Она тесно соединяется с железой соединительнотканными перемычками, продолжающимися в перегородки, отграничивающие друг от друга дольки железы. Глубокий листок капсулы в области глоточного отростка иногда отсутствует, что создает условия для распространения в окологлоточное пространство гнойного процесса при паротитах.
Околоушный проток (ductus parotideus), или Стенонов проток Название «Стенонов проток» образовано от имени анатома, его описавшего. Такие анатомические термины называются эпонимами. Эпонимы часто используются в клинической практике наряду с номенклатурными анатомическими терминами., формируется путем слияния междолевых протоков и достигает в диаметре 2 мм. Покидая железу у ее переднего края, он ложится на жевательную мышцу на 1 см ниже скуловой дуги, прободает щечную мышцу и открывается на слизистой оболочке щеки в преддверие рта на уровне 1-2-го верхних моляров. Добавочная околоушная железа, как правило, располагается над околоушным протоком, в который впадает ее собственный проток.
В толще околоушной железы проходит наружная сонная артерия и занижнечелюстная вена . Внутри железы наружная сонная артерия делится на две конечные ветви - верхнечелюстную и поверхностную височную артерии .
Сквозь околоушную железу проходит также лицевой нерв . В ней он делится на ряд ветвей, радиально расходящихся от области мочки уха к мимическим мышцам лица.
Кровоснабжение околоушной слюнной железы осуществляется ветвями наружной сонной артерии (a. carotis externa), среди которых задняя ушная артерия (a. auricularis posterior), проходящая косо назад над верхним краем заднего брюшка двубрюшной мышцы, поперечная артерия лица (a. transversa faciei) и скулоглазничная артерия (a. zygomaticoorbitalis), отходящая от поверхностной височной артерии (a. temporalis superficialis), а также глубокая ушная артерия (a. auricularis profunda), отходящая от верхнечелюстной артерии (a. maxillaris) (см. рис. 10). Выводной проток околоушной железы кровоснабжается из поперечной артерии лица. Артерии околоушной железы имеют многочисленные анастомозы между собой и с артериями близлежащих органов и тканей.
Венозный отток обеспечивается венами, сопровождающими выводные протоки железы. Сливаясь, они образуют вены околоушной жел езы (vv. parotideae), несущие кровь в занижнечелюстную (v. retromandibularis) и лицевую вены (v. facialis) и далее во внутреннюю яремную вену (v. jugularis interna).
На пути к занижнечелюстной вене кровь от верхней части железы оттекает также в поперечную вену лица (v. transversa faciei), от средней и нижней ее части - в жевательные вены (vv. maxillares) и крыловидное сплетение (plexus pterygoideus), от передней части железы - в передние ушные вены (vv. auriculares anteriores). От заушной части железы венозная кровь оттекает в заднюю ушную вену (v. auricularis posterior), иногда - в затылочные вены (vv. occipitales) и далее к наружной яремной вене (v. jugularis externa).
Лимфоотток осуществляется, в основном, в глубокие околоушные узлы (nodi parotidei profundi), в состав которых входят предушные, нижнеушные и внутрижелезистые узлы,
а также в поверхностные околоушные узлы (nodi parotidei superficiales). Из них лимфа направляется к поверхностным и латеральным глубоким шейным узлам .
Иннервация околоушной железы осуществляется околоушными ветвями ушно-височного нерва (n. auriculotemporalis), отходящего от нижнечелюстного нерва (n. mandibularis - III ветвь n. trigeminus). Околоушные ветви (rr. parotidei) включают чувствительные, следующие в составе тройничного нерва , и вегетативные нервные волокна.
Вегетативная иннервация околоушной железы осуществляется парасимпатическими постганглионарными нервными волокнами, отходящими от ушного узла (ganglion oticum), расположенного на медиальной поверхности нижнечелюстного нерва под овальным отверстием, и симпатическими постганглионарными нервными волокнами, отходящими от верхнего шейного узла (ganglion cervicale superius).
Преганглионарные парасимпатические нервные волокна берут свое начало от нижнего слюноотделительного ядра (nucl. salivatorius inf.), расположенного в продолговатом мозге; затем в составе языкоглоточного нерва (n. glossopharyngeus - IX пара черепных нервов) и его ветвей (n. tympanicus, n. petrosus minor) достигают ушного узла (ganglion oticum). От ушного узла постганглионарные нервные волокна следуют в околоушной железе по ветвям ушно-височного нерва .
Парасимпатические нервные волокна возбуждают секрецию железы и расширяют ее кровеносные сосуды.
Преганглионарные симпатические нервные волокна начинаются от вегетативных ядер верхних грудных сегментов спинного мозга и в составе симпатического ствола достигают верхнего шейного узла.
Симпатические постганглионарные нервные волокна идут от верхнего шейного узла и подходят к околоушной железе в составе сплетения наружной сонной артерии (plexus caroticus externus) по ветвям наружной сонной артерии, кровоснабжающим железу. Симпатическая иннервация оказывает суживающее влияние на кровеносные сосуды и тормозит секрецию железы.
Секреция слюны контролируется вегетативной нервной системой. К слюииым железам направляются парасимпатические и симпатические нервы, которые достигают их, следуя различными путями. Внутри желез аксоны различного происхождения располагаются в виде пучков.
Нервные волокна, идущие в строме желез совместно с сосудами, направляются к гладким миоцитам артериол, секреторным и миоэпи- телиальным клеткам коицевых отделов, а также клеткам вставочных и исчерченных отделов. Аксоны, утрачивая оболочку из шваннов- ских клеток, проникают сквозь базальную мембрану и располагаются между секреторными клетками концевых отделов, заканчиваясь терминальными варикозными расширениями, содержащими пузырьки и митохондрии (гиполеммальный нейроэффекторный контакт). Часть аксонов не проникают сквозь базальную мембрану, образуя варикозные расширения вблизи от секреторных клеток (эпилеммаль- ный нейроэффекторный контакт). Волокна, иннервирующие протоки, располагаются преимущественно вне эпителия. Кровеносные сосуды слюнных желез иннервированы симпатическими и парасимпатическими аксонами.
«Классические» нейромедиаторы (ацетилхолин в парасимпатических и норадреналин в симпатических аксонах) накапливаются в мелких пузырьках. Иммуногистохимически в нервных волокнах слюнных желез обнаружены разнообразные нейропептидные медиаторы, которые накапливаются в крупных пузырьках с плотным центром - вещество Р, пептид, связанный с кальцитониновым геном (ПСКГ), вазоактивный интестинальный пептид (ВИП), С-краевой пептид нейропептида Y (CPON), пептид гистидин-метионин (ПГМ).
Наиболее многочисленны волокна, содержащие ВИП, ПГМ, CPON. Они располагаются вокруг концевых отделов, проникая в них, оплетают выводные протоки, мелкие сосуды. Значительно реже встречаются волокна, содержащие ПСКГ и вещество Р. Предполагается, что пептидергические волокна участвуют в регуляции кровотока и секреции.
Обнаружены также афферентные волокна, которые наиболее многочисленны вокруг крупных протоков; их окончания проникают через базальную мембрану и располагаются среди эпителиальных клеток. Содержащие вещество P безмиелиновые и тонкие миелиновые волокна, несущие ноцицептивные сигналы, располагаются вокруг концевых отделов, кровеносных сосудов н выводных протоков.
Нервы оказывают на железистые клетки слюнных желез, как минимум, четыре вида воздействия: гидрокинетическое (мобилизация воды), протеокинетическое (секреция белка), синтетическое (усиление синтеза) и трофическое (поддержание нормальной структуры и функции). Помимо влияния на железистые клетки, нервная стимуляция вызывает сокращение миоэпителиальных клеток, а также изменения сосудистого русла (вазомоторный эффект).
Стимуляция парасимпатических нервных волокон приводит к секреции значительного объема водянистой слюны с низким содержанием белков и высокими концентрациями электролитов. Стимуляция симпатических нервных волокон вызывает секрецию небольшого количества вязкой слюны с высоким содержанием слизи.
Большинство исследователей указывают на то, что к моменту рождения слюнные железы сформированы неполностью; их диффе- ренцировка завершается в основном к 6 мес - 2 годам жизни, однако морфогенез продолжается до 16-20 лет. При этом может изменяться и характер вырабатываемого секрета: так, в околоушной железе в течение первых лет жизни продуцируется слизистый секрет, который лишь с 3-го года становится серозным. После рождения снижается синтез эпителиальными клетками лизоцима и лактоферрина, однако прогрессивно увеличивается выработка секреторного компонента. Одновременно в строме железы возрастает количество плазматических клеток, вырабатывающих преимущественно IgA.
После 40 лет впервые отмечаются явления возрастной инволюции желез. Этот процесс усиливается в пожилом и старческом возрасте, что проявляется изменениями как в концевых отделах, так и в выводных протоках. Железы, имеющие в молодости сравнительно мономорфное строение, с возрастом характеризуются прогрессирующей гетероморфией.
Концевые отделы с возрастом приобретают ббльшие различия в размерах, форме и тинкториальных свойствах. Размеры клеток концевых отделов и содержание в них секреторных гранул падают, а активность их лизосомального аппарата нарастает, что согласуется с часто выявляемыми картинами лизосомального разрушения гранул секрета - кринофагии. Относительный объем, занимаемый в крупных и мелких железах клетками концевых отделов, при старении снижается в 1,5-2 раза. Часть концевых отделов атрофируются и замещаются соединительной тканью, которая разрастается как между дольками, так и внутри долек. Редукции подвергаются преимущественно белковые концевые отделы; слизистые отделы, напротив, увеличиваются в объеме и накапливают секрет. В околоушной железе к 80 годам (как и в раннем детстве) обнаруживаются преимущественно слизистые клетки.
Онкоциты. В слюнных железах людей старше 30 лет часто обнаруживают особые эпителиальные клетки - онкоциты, которые редко выявляются в более молодом возрасте и присутствуют практически в 100 % желез у людей старше 70 лет. Эти клетки встречаются поодиночке или группами, часто в центре долек, как в концевых отделах, так и в исчерченных и вставочных протоках. Они характеризуются большими размерами, резко оксифильной зернистой цитоплазмой, везикулярным или пикнотическим ядром (встречаются также двуядерные клетки). На электронно-микроскопическом уровне отличительной особенностью онкоцитов является наличие в их ци-
топлазме огромного количества митохондрий, заполняющих ббльшую часть ее объема.
Функциональная роль онкоцитов в слюнных железах, а также в некоторых других органах (щитовидная и околощитовидная железы) не определена. Традиционный взгляд на онкоциты как на дегенеративно измененные элементы не согласуется с их ультраструктуриы- ми признаками и с их активным участием в обмене биогенных аминов. Происхождение этих клеток также остается предметом дискуссии. По мнению ряда авторов, они возникают непосредственно из клеток концевых отделов и выводных протоков вследствие их изменений. Возможно также, что они образуются в результате своеобразного изменения хода дифференцировки камбиальных элементов эпителия желез. Онкоциты слюнных желез могут давать особые опухоли органа - онкоцитомы.
Выводные протоки. Объем, занимаемый исчерченными отделами, при старении снижается, тогда как междольковые выводные протоки неравномерно расширяются, в них нередко обнаруживают скопления уплотненного материала. Последние обычно окрашиваются оксифильно, могут иметь слоистое строение и содержать соли кальция. Формирование таких мелких обызвествленных телец (конкрементов) не считается показателем патологических процессов в железах, однако образование крупных конкрементов (диаметром от нескольких миллиметров до нескольких сантиметров), вызывающих нарушения оттока слюны, служит ведущим признаком заболевания, называемого слюннокаменной болезнью, или сиалолитиазом.
Стромальный компонент при старении характеризуется увеличением содержания волокон (фиброзом). Основные изменения при этом обусловлены увеличением объема и более плотным расположением коллагеновых волокон, однако одновременно наблюдается и утолщение эластических волокон.
В междольковых прослойках нарастает количество адипоцитов, которые в дальнейшем могут появляться в дольках желез, замещая концевые отделы. Этот процесс наиболее выражен в околоушной железе. В последней, например, при старении до 50 % концевых отделов замещаются жировой тканью. Местами, часто по ходу выводных протоков и субэпителиально, выявляются скопления лимфоидной ткани. Указанные процессы происходят как в крупных, так и в мелких слюнных железах.
Секреторную функцию слюнных желез у животных изучают в острых и хронических опытах. Острый метод заключается во введении под наркозом в проток железы канюли, через которую выделяется слюна. Хронический (по Павлову) - хирургическим методом одна из протоков железы выводится на щеку (фистула) и к ней фиксируют воронку для сбора слюны (рис. 13.5). экспериментальные методы
РИС. 13.5.
дают возможность исследовать влияние различных факторов (пищевых, нервных, гуморальных) на секреторную функцию слюнных желез. У человека используют капсулу Лешли - Красногорского, которую фиксируют на слизистой оболочке щеки напротив протока железы.
Секреция слюны осуществляется слюнными железами рефлекторно.
Околоушные железы, крупнейшие среди слюнных желез, образуют серозный секрет, в состав которого входят белки и значительное количество воды; его количество составляет до 60 % слюны.
Подчелюстные и подъязычная железы продуцируют смешанный серозно-слизистый секрет, в состав которого входят белки и слизь - муцин, в количестве 25-30% и 10-15 % соответственно. Мелкие железы языка и ротовой полости секретируют преимущественно слизь - муцин.
В сутки слюнными железами образуется 0,8-2,0 л слюны, которая содержит воду, электролиты (по составу те же, что в плазме крови), белки, ферменты, муцин, защитные факторы (бактерицидные, бактериостатические), инсулиноподобный белок, паротин. pH слюны 6,0-7,4. Сухой остаток составляют неорганические и органические вещества.
■ Ферменты слюны представляют: альфа-амилаза, которая начинает гидролиз углеводов до дисахаридов: ДНК-азы и РНК-азы - расщепляют аминокислоты: "лингвальная" липаза - производится слюнными железами языка и начинает гидролиз липидов. Значительная группа ферментов (более 20) участвуют в гидролизе веществ, образующих зубной налет, и тем самым уменьшают зубные наслоения.
■ Муцин является гликопротеином, который защищает слизистую оболочку ротовой полости от механического повреждения и способствует образованию пищевого комка.
К защитных факторов слюны относятся:
1 Лизоцим (мурамидаза), который разрушает мембраны бактерий, а именно - разрывает 1-4 связи между N-ацетил-мураминовою кислотой и N- ацетилглюкозамина - двух основных мукопептидов, входящих в состав мембран бактерий. Лизоцим поступает в ротовую полость вместе со слюной крупных и мелких слюнных желез, с тканевым экссудатом десневой жидкости и из лейкоцитов, входящих в состав слюны. При высокой концентрации лизоцима в ротовой полости бактериальная флора становится неэффективной.
2 Секреторные IgA, в меньшей степени - IgG и IgM. Секреторные IgA вырабатываются слюнными железами, они более устойчивы к пищеварительным секретов, чем те, что находятся в плазме крови, в то время как IgM преимущественно является экссудатом жидкости выделяется деснами. IgA облегчает агрегацию микробов, образуя комплексы с белками поверхности эпителия, защищает его и способствует повышению фагоцитарной активности лейкоцитов.
3 Пероксидазы и тиоцианаты слюны действуют как антибактериальные ферменты.

РИС. 13.6.
4 Насыщение слюны солями кальция уменьшает декальцификация эмали.
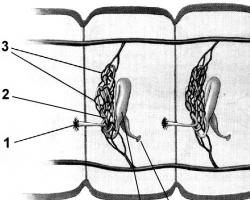
Механизм образования слюны , впервые описанный К. Людвигом, свидетельствует, что секреция не является пассивной фильтрацией жидкости из кровеносных сосудов - это результат активной функции секреторных клеток. В ацинарних клетках желез образуется первичная слюна. Ацинусни клетки синтезируют и выделяют ферменты и слизь, пролив - формируют жидкую часть слюны, ее ионный состав (рис. 13.6).
Фазы секреторного цикла. Через базальную мембрану капилляра в секреторную клетку проникают вещества, необходимые для синтеза ферментов, прежде всего - аминокислоты. Синтез просекрету (предшественника фермента) проходит на рибосомах, из которых он заносится в аппарат Гольджи на созревание. Зрелый секрет упаковывается в гранулы и сохраняется в них до времени выделения в просвет железы, который стимулируется ионами Са 2+.
Жидкая часть слюны образуется протоков клетками. Сначала она напоминает плазму крови, в которой высокая концентрация ионов натрия и хлора и значительно меньше - ионов калия и бикарбонатов. Образование жидкой слюны идет с затратами энергии с использованием кислорода, необходимого для синтеза АТФ. При прохождении слюны по протокам в ней меняется ионный состав - уменьшается количество натрия и хлора и увеличивается - ионов калия и бикарбонатов. Реабсорбция ионов натрия и секреция ионов калия регулируется альдостероном (как и в канальцах почек). В конечном итоге образуется вторичная слюна, выделяемая в ротовую полость (см. Рис. 13.6). На слиноутворення влияет уровень кровотока в железе, который зависит от образованных в ней метаболитов, особенно кининов (брадикинина), вызывающие местное расширение сосудов, увеличение секреции.
В ответ на действие различных раздражителей (с разными свойствами) слюнные железы выделяют неодинаковое количество слюны, с разным ее составом. Так, при употреблении сухой пищи выделяется большое количество жидкой слюны; при потреблении жидкой (молока) - производится мало, однако в ней много слизи.
Иннервация слюнных желез осуществляется парасимпатическими и симпатическими нервами. Парасимпатическую иннервацию железы получают от ядер черепных нервов продолговатого мозга: околоушные - от нижнего слюноотделительного ядра - IX пары (языко-глоточный), подчелюстные и подъязычная - от верхнего слюноотделительного ядра - VII пары (лицевой). Стимуляция парасимпатической нервной системы вызывает выделение большого количества жидкой слюны, бедной органическими веществами.
Симпатичную иннервацию всем слюнных желез дают центры боковых рогов II-IV грудных сегментов спинного мозга, через верхний шейный симпатический ганглий направляются к железам. При активации симпатических нервов слюны выделяется мало, но в ней высокая концентрация органических веществ (ферментов, муцина).
Регуляция слюноотделения осуществляется складнорефлекторнимы механизмами с помощью:
1 условных рефлексов на вид и запах пищи, звуки, сопровождающие акт еды их центр расположен в коре головного мозга (условнорефлекторная фаза) 2 безусловных рефлексов, связанных с раздражением пищей рецепторов языка, слизистой оболочки полости рта; их центр находится в слюноотделительного ядрах продолговатого мозга (безумовнорефлекторна фаза). Афферентный вход в ЦНС при реализации безусловных рефлексов - чувствительные волокна V, VII, IX и X пар черепных нервов; эфферентный выход - парасимпатические волокна VII, IX пар и симпатические нейроны боковых рогов II-IV сегментов грудного отдела (рис 13.7).
Пищеварение – включает комплекс механических и химических процессов, направленных на обработку пищи, всасывание питательных веществ, секрецию специальных ферментов в ротовой полости, желудке и кишечнике, выделение непереваренных компонентов пищи.
Внутриклеточное и пристеночное пищеварение. В зависимости от локализации процесса пищеварения его делят на внутриклеточное и внеклеточное. Внутриклеточное пищеварение - это гидролиз пищевых веществ, которые попадают внутрь клетки в результате фагоцитоза и пиноцитоза. В организме человека внутриклеточное пищеварение имеет место в лейкоцитах и в клетках лимфо-ретикуло-гистиоцитарной системы.
Внеклеточное пищеварение подразделяют на дистантное (полостное) и контактное (пристеночное, мембранное).
Дистантное (полостное) пищеварение осуществляется на значительном расстоянии от места образования ферментов. Ферменты в составе пищеварительных секретов осуществляют гидролиз пищевых веществ в полостях желудочно-кишечного тракта.
Контактное (пристеночное, мембранное) пищеварение осуществляется ферментами, фиксированными на клеточной мембране (А. М. Уголев). Структуры, на которых фиксированы ферменты, представлены в тонком отделе кишечника гликокаликсом. Первоначально гидролиз пищевых веществ начинается в просвете тонкой кишки под влиянием ферментов поджелудочной железы. Затем образовавшиеся олигомеры гидролизуются в зоне гликокаликса адсорбированными здесь ферментами поджелудочной железы. Непосредственно у мембран клеток кишечника гидролиз образовавшихся димеров производят фиксированные на ней собственно кишечные ферменты. Эти ферменты синтезируются в энтероцитах и переносятся на мембраны их микроворсинок.
Принципы регуляции процессов пищеварения . Деятельность пищеварительной системы регулируется нервными и гуморальными механизмами. Нервная регуляция пищеварительных функций осуществляется симпатическими и парасимпатическими влияниями.
Секреция пищеварительных желез осуществляется условно-рефлекторно и безусловно-рефлекторно. Такие влияния особенно выражены в верхней части пищеварительного тракта. По мере продвижения к дистальным отделам пищеварительного тракта участие рефлекторных механизмов в регуляции пищеварительных функций уменьшается. При этом повышается значение гуморальных механизмов. В тонком и толстом отделах кишечника особенно велика роль локальных механизмов регуляции - местное механическое и химическое раздражение повышает активность кишечника в месте действия раздражителя. Таким образом, в пищеварительном тракте существует градиент распределения нервных, гуморальных и местных регуляторных механизмов.
Местные механические и химические раздражители влияют на функции пищеварительного тракта посредством периферических рефлексов и через гормоны пищеварительного тракта. Химическими стимуляторами нервных окончаний в желудочно-кишечном тракте являются кислоты, щелочи и продукты гидролиза пищевых веществ. Поступая в кровь, эти вещества приносятся ее током к пищеварительным железам и возбуждают их непосредственно или через посредников. Объем крови, поступающий в желудок, кишечник, печень, поджелудочную железу и селезенку, составляет около 30% от ударного объема сердца.
Значительная роль в гуморальной регуляции деятельности органов пищеварения принадлежит гастроинтестинальным гормонам, образующимся в эндокринных клетках слизистой оболочки желудка, двенадцатиперстной кишки, тощей кишки, в поджелудочной железе. Они оказывают влияние на моторику пищеварительного тракта, на секрецию воды, электролитов и ферментов, на всасывание воды, электролитов и питательных веществ, на функциональную активность эндокринных клеток желудочно-кишечного тракта. Кроме того, гастроинтестинальные гормоны оказывают влияние на обмен веществ, эндокринные и сердечно-сосудистые функции, на центральную нервную систему. Некоторые гастроинтестинальные пептиды обнаружены в различных структурах мозга.
По характеру влияний регуляторные механизмы можно разделить на пусковые и корригирующие. Последние обеспечивают приспособление объема и состава пищеварительных соков к количеству и качеству пищевого содержимого желудка и кишечника (Г.Ф. Коротько).