Хроматин – комплекс ядерной ДНК с белками (гистоны, негистоновые белки).
Гетерохроматин (транкрипционно неактивный , конденсированный хроматин) интерфазного ядра. В СМ (видны в световой микроскоп)– базофильные глыбки, в ЭМ (видно в электронный микроскоп) – скопления плотных гранул. Располагается преимущественно по периферии ядра и вокруг ядрышек. Типичный пример гетерохроматина -тельце Барра .
Во всех соматических клетках генетически женского организма одна из Х –хромосом инактивирована и известна как половой хроматин (тельце Барра). Инактивация Х-хромосомы известна каклайонизация.
Лайонизация – механизм компенсации дозы генов Х-хромосомы у женщин объясняет гипотизаМэри Лайон.
Согласно гипотезе, инактивация Х-хромосомы происходит в раннем эмриогенезе, осуществляеися случайным образом (инактивированной может быть либо отцовская, либо материнская Х-хромосома), затрагивает целиком всю Х-хромосому и характеризуется устойчивостью, передаваясь клеточным потомкам. Клетки женского организма по экспрессии генов Х-хромосомы мозаичны.
Эухроматин – транскрипционно активная и менее конденсированная часть хроматина, локализуется в более светлых участках ядра между гетерохроматином.
Хромосомы видны при митозе или мейозе, когда хроматин конденсирован полностью.
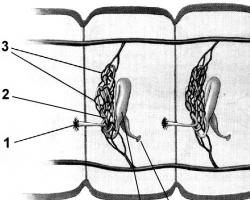
Рис. 8. Организация хроматина в хромосоме. Хроматин состоит из структурных единиц - нуклеосом, разделённых интервалами в 200 пар оснований. Во время митоза в результате плотной упаковки нуклеосом хроматин полностью конденсируется, формируя видимые хромосомы (из Widneil СС, Pfeninger КН, 1990)
ОРГАНИЗАЦИЯ ХРОМАТИНА В ХРОМОСОМЕ
Хроматин состоит из структурных единиц – нуклеосом, разделенных интервалами в 200 пар оснований. Во время митоза в результате плотной упаковки нуклеосом хроматин полностью конденсируется, формируя видимые хромосомы. (рис. 8)
Каждая хромосома содержит одну молекулу ДНК и ДНК- связывающие белки; хроматин в составе хромосомы образует многочисленные петли. Хромосома состоит из структурных единиц – нуклеосом. (рис. 9, 10)

Рис.9. Нуклеосома в неконденсированном хроматине содержит по две копии гистонов Н2А, Н2В, НЗ и Н4. Двойная спираль ДНК лежит на поверхности октамера гистонов и накручена на него. В конденсированном хроматине дополнительно присутствует гистон H 1, соединяющий нуклеосомы [из Trifonov EN , 1981|
Нуклеосомы – сферические структуры диаметром 10 нм.
 I
I
 III
III
 IV
IV
 V
V
Рис . 10. Уровни упаковки ДНК в хромосоме I – нуклеиновая нить,II – хроматиновая фибрилла,III – серия петельных доменов,IV – конденсированный хроматин в составе петельного домена,V – метафазная хромосома; 1 – гистон Н1, 2 – ДНК, 3 – прочие гистоны, 4 – микротрубочки ахроматинового веретена, 5 – кинетохор, 6 – центромера, 7 – хроматиды (по Б. Албертсу и соавт., с изменениями и дополнениями).
ГЕНОМ
Геном – полный комплект генов в хромосомах.Кариотип – описывает количество и структуру хромосом.Гаплоидный набор – 23 хромосомы – характерен для гамет.Диплоидный набор - стандарт хромосом (23 х 2) – для соматических клеток.
Соматических клеток самок плацентарных млекопитающих , включая человека. Хорошо прокрашивается осно́вными красителями .
Таким образом, у самки млекопитающего, гетерозиготной по какому-либо признаку, определяемому геном X-хромосомы, в разных клетках работают разные аллели этого гена (мозаицизм). Классическим видимым примером такого мозаицизма является окраска черепаховых кошек - в половине клеток активна X-хромосома с «рыжим», а в половине - с «чёрным» аллелем гена, участвующего в формировании меланина . Коты черепаховой окраски встречаются крайне редко и имеют две X-хромосомы (анеуплоидия) .
У людей и животных с анеуплоидией, имеющих в геноме 3 и более X-хромосом (см., напр., синдром Клайнфельтера), число телец Барра в ядре соматической клетки на единицу меньше числа X-хромосом.
Wikimedia Foundation . 2010 .
Тельце Барра. См. половой хроматин. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
тельце барра - ЭМБРИОЛОГИЯ ЖИВОТНЫХ ТЕЛЬЦЕ БАРРА – во всех соматических клетках генетически женского организма одна из Х хромосом инактивирована и известна как половой хроматин … Общая эмбриология: Терминологический словарь
Тельце Барра - интенсивно красящаяся структура, наблюдающаяся в ядрах разных типов клеток у самок млекопитающих. Представляет собой сильно спирализованную и поэтому неактивную Х хромосому. Инактивация одной из Х хромосом происходит случайным образом … Словарь по психогенетике
См. Хроматин половой. Источник: Медицинский словарь … Медицинские термины
- (М. L. Barr, род. в 1908 г., канадский гистолог) см. Хроматин половой … Большой медицинский словарь
Черепаховая окраска самок кошек является видимым примером инактивации Х хромосомы. Черный и оранжевый аллели гена окраски меха располагаются на Х хромосоме. Окраска конкретного участка меха определяется тем, какой из аллелей гена активен в данном … Википедия
половой хроматин - тельце Барра Гетерохроматинизированная в процессе лайонизации и интенсивно прокрашиваемая при анализе интерфазных ядер Х хромосома самок млекопитающих; выявление П.х. лежит в основе экспресс метода (т.е. без культивирования клеток и получения… … Справочник технического переводчика
Плотное окрашивающееся тельце, обнаруживаемое в недслящихся (интерфазных см. Интерфаза) ядрах клеток у гетерогаметных (имеющих Х и Y Половые хромосомы) животных и человека. П. х. подразделяют на Х хроматин, или тельце Барра (открыт в 1949 … Большая советская энциклопедия
Ядро фибробласта женщины с гетерохроматином, помеченным флуоресцентным красителем. Стрелкой указано положение тельца Барра Гетерохроматин участки … Википедия
I совокупность генетически детерминированных признаков особи, определяющих ее роль в процессе размножения. Развитие признаков мужского (обозначают знакома Марса ♂) и женского (знак Венеры ♀) полов определяется хромосомными наборами (см.… … Медицинская энциклопедия
В 1949 г. М. Барр и Ч. Бертрам, изучая нейроны кошки, обратили внимание на то, что в интерфазном ядре клетки содержится интенсивно окрашиваемое тельце, причем оно присутствует только в ядрах клеток самок и отсутствует у самцов. Оно было найдено у многих животных и всегда только у особей женского пола. Это тельце получило название полового хроматина, или тельца Барра. У ряда позвоночных и у человека оно появляется в раннем онтогенезе на стадии гаструлы, но раньше развития гонад (половых желез). На локализацию, форму и структуру полового хроматина не влияют половые гормоны, следовательно, он не является вторичным половым признаком. Между числом телец полового хроматина и числом X- хромосом в ядре имеется прямая связь. Половой хроматин в интерфазных ядрах обусловлен спирализацией одной из Х-хромосом, инактивация которой является механизмом, выравнивающим баланс генов половых хромосом в клетках самцов и самок (т. е. это один из механизмов дозовой компенсации генов). 6
В 1961 г. несколько исследователей одновременно высказали предположения, что одна из Х-хромосом у нормальных женщин относительно не активна в генетическом отношении. В 1961 году английская исследовательница М. Лайон выдвинула гипотезу о механизмах инактивации одной из Х-хромосом клеток женского организма. Основные положения этой гипотезы следующие:
1. Одна из двух Х-хромосом клеток женщины неактивна.
2. Неактивная хромосома может быть отцовского или материнского организма.
3. Инактивация происходит в раннем эмбриогенезе и сохраняется во время дальнейшего размножения и развития клеточной линии. Этот процесс инактивации Х-хромосомы в ряду поколений обратим:
XX* ->- УХ -> XX* и т. д. (здесь звездочкой обозначена спирали-зованная Х-хромосома). Такой тип обратимых изменений генетического материала португальский генетик Серра предложил называть трепцией (от греч. treptos - изменение).
Спирализованная Х-хромосома в клетке образует половой хроматин или тельце Барра. Если у женщин в ядре клетки несколько Х-хромосом, то в клетках несколько телец Барра, активной остается лишь одна Х-хромосома. Х-хромосома инактивируется не вся, часть короткого плеча остается генетически активной. Инактивация Х-хромосомы в определенной мере зависит от стадии клеточного цикла и физиологического состояния организма. По наличию лишнего или отсутствию тельца Барра можно диагносцировать некоторые виды наследственных заболеваний (например, синдром Клайнфельтера, синдром Шерешевского - Тернера). Клетки, не содержащие половой хроматин (хроматин-отрицательные клетки), обнаруживаются у индивидуумов, имеющих набор хромосом 45, ХО (синдром Шерешевского - Тернера);
46, XY (нормальные мужчины); 47, XYY (синдром Клайнфельтера с двумя Y-хромосомами). Обычно в клетках нормального мужского организма встречается некоторое количество псевдотелец Барра (конденсированных участков аутосом) и спирализованных Y-хромосом, поэтому при диагностике различных хромосомных заболеваний необходимо уметь отличать эти образования от типичного полового хроматина, образованного спирализованной лишней Х-хромосомой. Тельце Барра обнаруживается при хромосомном наборе 46, XX (нормальные женщины); 47, ХХУ и 48, ХХУУ (классический синдром Клайнфельтера). Два тельца Барра обнаруживаются у человека, имеющего три Х-хромосомы, (47, XXX); три Х-хромосомы и одну У (48, ХХХУ, синдром Клайнфельтера); 49, ХХХУУ (синдром Клайнфельтера). Три тельца Барра встречаются при кариотипе 48, ХХХХ и 49, ХХХХУ (тяжелый синдром Клайнфельтера).
В полиплоидных клетках число телец полового хроматина соответствует плоидности. По формуле Гарднера, число телец Барра (В)
равно В = Х - , где Х - число Х-хромосом, Р - степень плоидности клетки. В неполиплоидных клетках число телец полового хроматина равно числу Х-хромосом минус единица (В = Х - 1).
Структурные изменения хромосом
Хромосомы могут подвергаться различным структурным изменениям. Особенно важное значение имеют потеря отдельных фрагментов хромосом (деления) или перенос участка одной хромосомы на другую (транслокация). Транслокация обозначается латинской буквой /, в скобках рядом с ней пишут индекс группы или номер хромосомы-донора, обозначение переносимого участка. Эти же обозначения указываются для хромосомы-реципиента, например 46, XXt (Ср + + В4q -). В скобках буквами р и q указывают плечи хромосом, затрагиваемые транслокацией. Короткое плечо хромосомы обозначают буквой р, длинное - буквой q, спутник - буквой s, и т. д. Увеличение длины плеча обозначается знаком плюс, а уменьшение - знаком минус (оба они ставятся после символа хромосомы).
Появление одной лишней хромосомы в кариотипе приводит к трисомии. Кратное увеличение числа всех хромосом носит название полиплоидии (могут быть триплоиды, тетраплоиды и т. д.). Потеря одной из пары гомологичных хромосом приводит к состоянию, которое называется моносомией. Изменения числа или строения хромосом называется хромосомными аберрациями.
Рассмотрим наиболее частые виды структурных нарушений хромосом - делеции и транслокации. При делеции общее количество хромосом не изменено. Однако в какой-то хромосоме недостает генетического материала, что вызывает различные изменения фенотипа. Чаще всего встречается делеция 5-й и 18-й аутосом и Х-хромосомы. Делеции приводят к развитию различных наследственных заболеваний и синдромов.
В 1963 г. Ж. Лежен описал синдром «кошачьего крика». Крик таких детей напоминает «мяуканье кошки». У детей резкое недоразвитие гортани, круглое лунообразное лицо, микроцефалия, микрогнатия, монголоидный разрез глаз, низко расположенные деформированные ушные раковины, мышечная гипотония, слабо выраженные вторичные половые признаки. Эти дети умственно отсталые. В кариотипе детей отмечается делеция короткого плеча 5-й пары хромосом.
Деления длинного и короткого плеча 18-й хромосомы сопровождается различными нарушениями строения лица, скелета, внутренних органов. У детей отмечается умственная отсталость, гипотрофия, гипотония, микроцефалия, недоразвитие лица, низкий грубый голос, недоразвитие наружных половых органов, среднего уха, атрезия наружного слухового прохода и другие аномалии.
При делеции короткого плеча 18-й хромосомы у больных также отмечаются различные дефекты со стороны скелета, внутренних органов и умственная отсталость.
Делеция короткого плеча Х-хромосомы может трактоваться как частичная моносомия по Х-хромосоме. Описана у женщин, у которых наблюдается задержка роста, недоразвитие яичников без тяжелых соматических аномалий. Хотя половой хроматин у них выявляется, однако его размеры значительно меньше, чем в норме.
При хронических миелолейкозах отмечается укорочение короткого плеча 21-й хромосомы (так называемая филадельфийская хромосома). Однако эта хромосома обнаруживается только в клетках крови и пунктате костного мозга. Другие же клетки имеют нормальный кариотип.
В результате двух концевых нехваток с последующим соединением разорванных концов образуются кольцевые хромосомы. Поэтому данное нарушение структуры хромосом фактически является частным случаем делеции. Клиническая картина больных - носителей кольцевых хромосом - напоминает таковую при делеции соответствующей хромосомы. Так, при кольцевой хромосоме группы В (5-я пара) развивается клиническая картина синдрома «кошачьего крика», а при кольцевой Х-хромосоме клиническая картина близка синдрому Шерешевского - Тернера.
Транслокации - это структурные перестройки, при которых происходит обмен генетического материала между хромосомами. Возможны различные виды транслокаций: реципрокные, при которых происходит взаимный обмен фрагментами; нереципрокные, когда генетический материал одной хромосомы переносится на другую, и наконец центрические соединения. Наиболее часто встречаются именно последние транслокации между акроцентрическими хромосомами. При этом утрачивается только небольшой фрагмент коротких плечей акроцентрических хромосом. Большую часть таких перестроек можно считать сбалансированной, так как они не вызывают серьезных отклонений в фенотипе носителя транслокации. Однако потомство таких носителей имеет клинически выраженные дефекты, характерные для аномального набора хромосом.
Известно, что болезнь Дауна может наблюдаться как при трисомии по 21-й аутосоме, так и при транслокации фрагмента этой хромосомы на другие. У таких больных хромосом 46, но одна из хромосом фактически двойная, так как к ней еще прикреплен фрагмент 21-й хромосомы и в результате такая перестройка оказывается не сбалансированной. У родителей этих больных кариотип включал 45 хромосом, но одна из хромосом была фактически двойной (с транслокацией). При оплодотворении яйцеклетки, содержащей эту хромосому, нормальным спермием в зиготе фактически будут три 21-х хромосомы, что фенотипически проявляется болезнью Дауна.
21-я хромосома чаще всего транслоцируется на 15-ю или на другие хромосомы группы Д (13-ю, 14-ю) у женщин, или на 22-ю у мужчин. В таком случае у молодых здоровых родителей может родиться ребенок с болезнью Дауна в отличие от трисомии 21-й хромосомы, которая чаще бывает у детей, рожденных пожилыми матерями. Определить наличие транслокации у индивидуума до рождения ребенка с болезнью Дауна без исследования кариотипа фактически невозможно, так как фенотип этих носителей мало чем отличается от фенотипов лиц с нормальными генотипами. Поэтому во всех этих случаях исследование кариотипа имеет особенно важное значение.
Механизм развития болезни Дауна при транслокации у одного из родителей можно представить следующим образом. При транслокации кариотип индивидуума состоит из 45 хромосом, так как одна хромосома увеличена в размере. Транслокация касается всех клеток, в том числе и оогоний и сперматогоний. При образовании половых клеток (гамет) в одну гамету попадает 23 хромосомы, а в другую 22. Но транслоцированная хромосома может оказаться как в гамете с 22 хромосомами, так и в гамете с 23 хромосомами. Таким образом, теоретически возможны 4 варианта гамет: 23 нормальные хромосомы, 23 с транслокацией, 22 нормальные хромосомы и 22 с транслокацией. Если транслокацию обозначить апострофом, то получится следующий ряд гамет: 23 23 1 22 22 1 .
Если эти гаметы будут оплодотворены нормальной гаметой противоположного пола, то получим следующие комбинации: 1) 23 + 23 = = 46 хромосом (нормальный кариотип); 2) 23 1 + 23 = 46 1 хромосом, но фактически 47 хромосом (в данном случае разовьется болезнь Дауна); 3) 22 + 23 = 45 хромосом (такая зигота не жизнеспособна и погибает); 4) 22 1 +23 = 45 1 хромосом (в этом случае рождается индивидуум с транслокацией, как и один из его родителей).
Шансы родить ребенка с болезнью Дауна (при транслокации у одного из родителей) составляют 33%. Это очень большой риск и в таком случае дальнейшее деторождение не желательно, тем более что есть риск получить транслокацию и у внуков. Если рождается ребенок с болезнью Дауна, вызванной трисомией по 21-й хромосоме, у родителей с нормальным кариотипом, то шансы родить повторно такого же ребенка очень незначительны. Однако не во всех случаях при рождении ребенка с болезнью Дауна вследствие транслокации 21-й хромосомы транслокация имеется в соматических клетках матери. Примерно у половины матерей кариотип бывает нормальный, а транслокация произошла во время мейоза, предшествующего образованию яйцеклетки, из которой развился организм больного ребенка.
Половые хромосомы (гоносомы, гетеросомы) различаются как по строению (длина, положение центромеры, количество гетерохроматина), так и по содержанию генов.
Хромосома X - это субметацентрическая хромосома средних размеров, входит в группу С). Она есть в соматических клетках индивидов обоих полов: в двойном экземпляре у женщин с кариотипом 46,ХХ и в одном экземпляре у мужчин с кариотипом 46,ХY; а также в одном экземпляре во всех яйцеклетках и 50% сперматозоидов.Хромосома X богата эухроматиновыми участками и содержит 1336 генов, среди которых: соматические гены, регуляторные гены феминизации, структурные гены феминизации, структурные гены маскулинизации. Таким образом, хромосома Х является обязательной в кариотипе соматической клетки как женского, так и мужского полов.
Хромосома Y является мелкой акроцентрической хромосомой, входит в группу G; 2/3 дистального плеча q представлены гетерохроматином и генетически неактивны. Хромосома Y представлена одним экземпляром во всех соматических клетках индивидов мужского пола с кариотипом 46,XY и у 50% сперматозоидов. Она содержит 307 генов, среди которых: регуляторные гены маскулинизации (SRY + Tdf), гены, обеспечивающие фертильность (AZF1, AZF2), несколько структурных соматических генов и псевдогены.
Морфологические и генетические различия между хромосомами X и Y, а также отличия по количеству хромосом Х в кариотипе стало причиной генетического неравенства между полами (у женщин по сравнению с мужчинами двойная доза генов хромосомы Х. Однако это неравенство не проявляется, благодаря механизму компенсации, в результате которого функциональной остается только одна хромосома Х в соматических клетках и мужчин и женщин, а именно:
В клетках 46,ХХ – активна только одна хромосома Х;
В клетках 46,XY – активны хромосомы Х и Y;
В клетках 47,ХХХ – активна только одна хромосома Х;
В клетках 47,ХХY – активна только одна хромосома Х и одна хромосома Y;
В клетках 48,ХХХY – активна только одна хромосома Х и одна Y;
Путем гетерохроматинизации одной из двух хромосом Х и женщин образуется половой хроматин Х, а в результате гетерохроматинизации 2/3q хромосомы Y у лиц мужского пола образуется половой хроматин Y.
Половой хроматин Х:
Представляет инактивированную хромосому Х. в форме факультативного гетерохроматина, в соматических клетках 46,ХХ;
Выявляется в интерфазных ядрах соматических клеток в виде тельца Барра размером около 1µm;
Тест Барра используется для определения количества хромосом Х в кариотипе в норме и в случае гносомных анеуплоидий;
Число хр.Х = числу телец Барра + 1 (активная хр.Х);
46,ХХ – 1 тельце Барра;
46,ХY – отсутствует тельце Барра;
47,ХХХ – 2 тельца Барра;
47,ХХY – 1 тельце Барра;
45,Х – отсутствует тельце Барра;
48,ХХХХ – 3 тельца Барра.
Половой хроматин Y:
Представлен 2/3 плеча Y q хромосомы Y, в форме конститутивного гетерохроматина, в соматических клетках 46,XY и сперматозоидах 23,Y;
Выявляется в интерфазных ядрах клеток в виде тельца F (флуоресцентного) размером около 1µm;
Тест F используется для идентификации хромосомы Y (пренатальное определение пола);
Число хр.Y = числу телец F;
46,ХХ – отсутствует тельце F;
46,ХY – 1 тельце F;
47,ХYY – 2 тельца F;
47,ХХY – 1 тельце F;
48,ХХYY – 2 тельца F;
46,X,i(Yp) – отсутствует тельце F;
46,X,i(Yq) – 1(0,5 µm) тельце F.