Мембранный потенциал покоя (МПС) — это разность потенциалов между внешней и внутренней сторонами мембраны в условиях, когда клетка не возбуждено. Цитоплазма клетки заряжена отрицательно к внеклеточной жидкости неравномерным распределением анионов и катионов по обе стороны мембраны. Разность потенциалов (напряжение) для различных клеток имеет значение от -50 до -200 мВ (минус означает, что внутри клетка более негативно заряжена, чем снаружи). Мембранный потенциал покоя возникает на мембранах всех клеток — возбуждающих (нервов, мышц, секреторных клеток) и незбудливих.
МПС необходим для поддержания возбудимости таких клеток, как мышечные и нервовои. Также он влияет на транспорт всех заряженных частиц в любом типе клеток: он способствует пассивному транспорта анионов из клетки и катионов в клетку.
Образование и поддержания мембранного потенциала обеспечивают различные типы ионных насосов (в частности натрий-калиевый насос или натрий-калиевая АТФаза) и ионных каналов (калиевые, натриевые, хлорные ионные каналы).
Для регистрации потенциала покоя используют специальную микроэлектродную технику. Микроэлектрод — это тоненькая стеклянная трубочка, с вытянутым концом, диаметром менее 1 мкм, заполненная раствором электролита (чаще хлорида калия). Рефернтним электродом служит серебряная хлорированная пластинка, расположенная в внеклеточном пространстве, оба электрода подключены к осциллографа. Сначала оба электрода занходяться в внеклеточном пространстве и разность потенциалов между ними отсутствует, если ввести регистрирующий микроэлектрод через мембрану в клетку, то осциллограф покажет скачкообразное смещение потенциала примерно до -80 мВ. Этот сдвиг потенциала называют мембранным потенциалом покоя.
К возникновению мембранного потенциала покоя приводят два фактора: во-первых, концентрации различных ионов отличаются внешне и всереднини клетки, во-вторых мембрана является полупроницаемой: одни ионы могут через нее проникать, другие — нет. Оба эти явления зависят от наличия в мембране специальных белков: концентрационные градиенты создают ионные насосы, а проницаемость мембраны для ионов обеспечивают ионные каналы. Важнейшую роль в формировании мембранного потенциала играют ионы калия, натрия и хлора. Концентрации этих ионов видризняюються по обе стороны мембраны. Для нейрона млекопитающих концентрация K + составляет 140 ммоль внутри клетки и только 5 мМ извне, градиент концентрации Na + почти противоположный — 150 ммоль снаружи и 15 мМ внутри. Такое распределение ионов поддерживается натрий-калиевым насосом в плазматической мембране — белком использующий энергию АТФ для закачки K + в клетку и скачивания Na + из нее. Также существует концентрационный градиент и для других ионов, например, хлорид аниона Cl -.
Концентрационные градиенты катионов калия и натрия — это химическая форма потенциальной энергии. В преобразовании энергии в электрическую участвуют ионные каналы — поры формируются скоплениями специальных трансмембранных белков. Когда ионы диффундируют через канал, они переносят единицу электрического заряда. Любой суммарный движение положительных или отрицательных ионов через мембрану будет создавать напряжение, или разность потенциалов по обе стороны мембраны.
Ионные каналы, участвующие в утовренни МПС имеют избирательную проницаемость, то есть дают возможность проникать только определенному типу ионов. В мембране нейрона в состоянии покоя открытые калиевые каналы (те, что в основном пропускают только калий), большинство натриевых каналов — закрыты. Диффузия ионов K + через калиевые каналы является решающим для создания мембранного потенциала. Так как концентрация K + значительно выше внутри клетки, химический градиент способствует оттоку этих катионов из клетки, поэтому в цитоплазме начинают преобладать анионы, которые не могут проходить через калиевые каналы.
Отток ионов калия из клетки ограничен самым мембранным потенциалом, поскольку при определенном его уровне накопление отрицательных зарядов в цитоплазме будет ограничивать движение катионов за пределы клетки. Таким образом, главным фактором в возникновении МПС является распределение ионов калия под действием электрического и химического потенциалов.
Для того, чтобы определить влияние движения определенного иона через полупроницаемую мембрану на формирование мембранного потенциала, строят модельные системы. Такая модельная система состоит из сосуда разделенной на две ячейки искусственной полупроницаемой мембраной, в которую встроены ионные каналы. В каждую ячейку можно погрузить электрод и померить разность потенциалов.
Рассмотрим случай, когда искусственная мембрана проницаема только для калия. По две стороны мембраны модельной системы создают градиент концентрации аналогичный таковому у нейроне: в ячейку, соответствующую цитоплазме (внутренняя ячейка), помещают 140 мМ раствор хлорида калия (KCl), в ячейку, соответствующую межклеточной жидкости (внешняя ячейка) — 5 ммоль раствор KCl. Ионы калия будут диффундировать через мембрану во внешнюю ячейку по градиенту концентрации. Но поскольку анионы Cl — проникать через мембрану не могут во внутренней ячейке возникать избыток отрицательного заряда, который будет препятствовать подали оттока катионов. Когда такие модельные нейроны достигнут состояния равновесия, действие химического и электрического потенциала будет сбалансирована, ни суммарной диффузии К + не будет наблюдаться. Значение мембранного потенциала, виинкае при таких условиях, называется равновесным потенциалом для определенного иона (Е ион). Равновесный потенциал для калия составляет примерно -90 мВ.
Аналогичный опыт можно провести и для натрия, установив между ячейками мембрану проникающей только для этого катиона, и поместив во внешнюю ячейку раствор хлорида натрия с концентрацией 150 мМ, а во внутреннюю — 15 мМ. Натрий будет двигаться во внутреннюю ячейку, ривоноважний потенциал для него составит примерно 62 мВ.
Количество ионов, должна диффундировать для генерации электрического потенциала очень невелика (примерно 10 -12 моль К + на 1 см 2 мембраны), этот факт имеет два важных последствия. Во-прешь, это означает, что концентрации ионов, которые могут проникать через мембрану, остаются стабильными снаружи и внутри клетки, даже после того как их движение обеспечил утоврення электрического потенциала. Во-вторых, мизерные потоки ионов через мембрану, потирбно для установления потенциала, не нарушают электронейтральности цитоплазмы и внеклеточной жидкости в целом, распределение зарядов происходит только в области, непосредственно прилегающей к плазматической мембраны.
Равновесный потенциал для определенного иона, например для калия, можно рассчитать по уравнению Нернста, что выглядит так:
,где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, o, i — концентрация калия снаружи и внутри клетки соответственно. Поскольку описанные процессы происходят при температуре тела — 310 ° К, а десятичными логарифмами в исчислении пользоваться легче чем натуральными, это уравнение превращают следующим образом:
Подставляя концентрации К + в уравнение Нернста получаем равновесный потенциал для калия, составляет -90 мВ. Поскольку по нулевой потенциал принимается внешняя сторона мембраны, то знак минус означает, что в условиях равновесного калиевого потенциала внутренняя Сторн мембраны сравнительно более электроотрицательным. Аналогичные расчеты можно провести и для равновесного Натиева потенциала, он составляет 62 мВ.
Хотя равновесный потенциал для ионов калия составляет -90 мВ, МПС нейрона несколько менее отрицательный. Эта разница отражает незначительное но постоянное следование ионов Na + через мембрану в состоянии покоя. Поскольку концентрационный градиент для натрия противоположный такого для калия, Na + движется внутрь клетки и сдвигает суммарный заряд на внутренней стороне мембраны в положительную сторону. На самом деле МПС нейрона составляет от -60 до -80 мВ. Это значение значительно ближе к Е K чем до Е Na, потому что в состоянии покоя в нейроне открыто много калиевых каналов и очень мало натриевых. Также на встанвлення МПС влияет движение ионов хлора. В 1943 году Дэвид Голдаман предложил усовершенствовать уравнение Нернста так, чтобы оно отражало влияние различных ионов на мембарнний потенциал, в этом уравнении учитывается относительная проницаемость мембраны для каждого типа ионов:
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, [ион] o, [ион] i — концентрации ионов внутри и внутри клеток, Р — относительная проницаемость мембраны для соответствующего иона. Значение заряда в данном уравнении не сохраняется, но оно учтено — для хлора внешняя и внутренняя концентрация поменяны местами, так как его заряд 1.
Мембранный потенциал покоя может существовать только при условии неравномерного распределения ионов, обеспечивается функционированием натрий-калиевого насоса. Кроме того, этот белок делает также электрогенных властовости — он переносит 3 катионы натрия в обмен на 2 ионы калия, движущихся внутрь клетки. Таким образом, Na + -K + -АТФазы снижает МПС на 5-10 мВ. Подавление деятельности этого белка приводит к незначительному (на 5-10 мВ) мгновенного повышения мембранного потенциала, после чего он еще некоторое время будет существовать на достаточно стабильном уровне, пока будут сохраняться градиенты концентраций Na + и K +. Впоследствии эти градиенты начнут уменьшаться, вследствие проникнсоти мембраны к ионам, и через несколько десятков минут электрический потенциал на мембране исчезнет.
Установлено, что наиболее важными ионами, определяющими мембранные потенциалы клеток, являются неорганические ионы К + , Na + , СГ, а также в ряде случаев Са 2 + . Хорошо известно, что концентрации этих ионов в цитоплазме и в межклеточной жидкости различаются в десятки раз.
Из табл. 11.1 видно, что концентрация ионов К + внутри клетки в 40-60 раз выше, чем в межклеточной жидкости, тогда как для Na + и СГ распределение концентраций противоположное. Неравномерное распределение концентраций этих ионов по обе стороны мембраны обеспечивается как их различной проницаемостью, так и сильным электрическим полем мембраны, которое определяется ее потенциалом покоя.
Действительно, в состоянии покоя суммарный поток ионов через мембрану равен нулю, и тогда из уравнения Не- рнста - Планка следует, что
Таким образом, в покое градиенты концентрации - и
электрического потенциала -- на мембране направлены
противоположно друг другу и поэтому в покоящейся клетке высокая и постоянная разность концентраций основных ионов обеспечивает поддержание на мембране клетки электрического напряжения, которое и называют равновесным мембранным потенциалом.
В свою очередь возникающий на мембране потенциал покоя препятствует выходу ионов из клетки К + и чрезмерному входу в нее СГ, поддерживая тем самым их концентрационные градиенты на мембране.
Полное выражение для мембранного потенциала, учитывающее потоки диффузии этих трех видов ионов, было получено Гольдманом, Ходжкиным и Катцем:
где Р к, P Na , Р С1 - проницаемость мембраны для соответствующих ионов.
Уравнение (11.3) с высокой точностью определяет мембранные потенциалы покоя различных клеток. Из него следует, что для мембранного потенциала покоя важны не абсолютные величины проницаемостей мембраны для различных ионов, а их отношения, так как, разделив обе части дроби под знаком логарифма, например, на Р к, мы перейдем к относительным проницаемостям ионов.
В тех случаях, когда проницаемость одного из этих ионов значительно больше, чем других, уравнение (11.3) переходит в уравнение Нернста (11.1) для этого иона.
Из табл. 11.1 видно, что мембранный потенциал покоя клеток близок к потенциалу Нернста для ионов К + и СВ, но значительно отличается от него по Na + . Это свидетельствует
0 том, что в покое мембрана хорошо проницаема для ионов К + и СГ, тогда как для ионов Na + ее проницаемость очень низка.
Несмотря на то что равновесный потенциал Нернста для СГ наиболее близок к потенциалу покоя клетки, последний имеет преимущественно калиевую природу. Это обусловлено тем, что высокая внутриклеточная концентрация К + не может существенно уменьшиться, так как ионы К + должны уравновешивать внутри клетки объемный отрицательный заряд анионов. Внутриклеточные анионы представляют собой в основном крупные органические молекулы (белки, остатки органических кислот ит.п.), которые не могут пройти через каналы в клеточной мембране. Концентрация этих анионов в клетке практически постоянна и их суммарный отрицательный заряд препятствует значительному выходу калия из клетки, поддерживая вместе с Na-K-насосом его высокую внутриклеточную концентрацию . Однако основная роль в первоначальном установлении внутри клетки высокой концентрации ионов калия и низкой концентрации ионов натрия принадлежит Na-K-насосу.
Распределение ионов С1 устанавливается в соответствии с мембранным потенциалом, поскольку в клетке нет специальных механизмов поддержания концентрации СГ. Поэтому вследствие отрицательного заряда хлора его распределение оказывается обратным по отношению к распределению калия на мембране (см. табл. 11.1). Таким образом, концентрационные диффузии К + из клетки и С1 в клетку практически уравновешиваются мембранным потенциалом покоя клетки.
Что касается Na + , то в покое его диффузия направлена в клетку под действием как градиента концентрации, так и электрического поля мембраны и вход Na + в клетку ограничивается в покое только малой проницаемостью мембраны для натрия (закрыты натриевые каналы). Действительно, Ходжкин и Катц экспериментально установили, что в состоянии покоя проницаемости мембраны аксона кальмара для К + , Na + и СГ относятся как 1: 0,04: 0,45. Таким образом, в состоянии покоя клеточная мембрана малопроницаема только для Na + , а для СГ она проницаема почти так же хорошо, как и для К + . В нервных клетках проницаемость для СГ обычно ниже, чем для К + , но в мышечных волокнах проницаемость для СГ даже несколько преобладает.
Несмотря на малую проницаемость клеточной мембраны для Na + в покое, существует, хотя и весьма малый, пассивный перенос Na + в клетку. Этот ток Na + должен был бы приводить к снижению разности потенциалов на мембране и к выходу К + из клетки, что вело бы в конечном итоге к выравниванию концентраций Na + и К + по обе стороны мембраны. Этого не происходит благодаря работе Na + - К + -насоса, компенсирующего токи утечки Na + и К + и поддерживающего таким образом нормальные значения внутриклеточных концентраций этих ионов и, следовательно, нормальную величину потенциала покоя клетки.
Для большинства клеток мембранный потенциал покоя составляет (-бО)-(-ЮО) мВ. На первый взгляд может показаться, что это малая величина, но надо учесть, что толщина мембраны тоже мала (8-10 нм), так что напряженность электрического поля в клеточной мембране огромна и составляет около 10 млн вольт на 1 м (или 100 кВ на 1 см):
Воздух, например, не выдерживает такой напряженности электрического поля (электрический пробой в воздухе наступает при 30 кВ/см), а мембрана выдерживает. Это нормальное условие ее деятельности, поскольку именно такое электрическое поле необходимо для поддержания разности концентраций ионов натрия, калия и хлора на мембране.
Величина потенциала покоя, различная у клеток, может изменяться при изменении условий их жизнедеятельности. Так, нарушение биоэнергетических процессов в клетке, сопровождающееся падением внутриклеточного уровня макро- эргических соединений (в частности, АТФ), прежде всего исключает компоненту потенциала покоя, связанную с работой Ма + -К + -АТФ-азы.
Повреждение клетки приводит обычно к повышению проницаемости клеточных мембран, в результате чего различия в проницаемости мембраны для ионов калия и натрия уменьшаются; потенциал покоя при этом уменьшается, что может вызвать нарушение ряда функций клетки, например возбудимости.
Все живые клетки обладают способностью под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности или возбуждения.
Возбуждение - это комплекс активных электрических, химических и функциональных изменений в возбудимых тканях (нервной, мышечной или железистой), которыми ткань отвечает на внешнее воздействие. Важную роль при возбуждении играют электрические процессы, обеспечивающие проведение возбуждения по нервным волокнам и приводящие ткани в активное (рабочее) состояние.
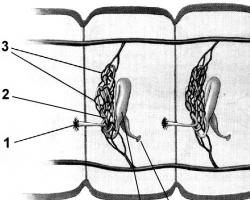
Живые клетки обладают важным свойством: внутренняя поверхность клетки всегда заряжена отрицательно по отношению к внешней ее стороне. Между внешней поверхностью клетки, заряженной электроположительно по отношению к протоплазме, и внутренней стороной клеточной мембраны существует разность потенциалов, которая колеблется в пределах 60-70 мВ. По данным П. Г. Ко- стюка (2001), у нервной клетки эта разность колеблется в пределах от 30 до 70 мВ. Разность потенциалов между внешней и внутренней сторонами мембраны клетки называют мембранным потенциалом, или потенциалом покоя (рис. 2.1).
Мембранный потенциал покоя присутствует на мембране до тех пор, пока клетка жива, и исчезает с гибелью клетки. Л. Гальвани еще в 1794 г. показал, что если повредить нерв или мышцу, сделав поперечное сечение и приложив к поврежденной части и к месту повреждения электроды, соединенные с гальванометром, то гальванометр покажет ток, который течет всегда от неповрежденной части ткани к месту разреза. Это течение он назвал током покоя. По своей физиологической сути ток покоя и мембранный потенциал покоя - одно и то же. Измеренная в данном опыте разность потенциалов составляет 30-50 мВ, поскольку при повреждении ткани часть тока шунтируется в межклеточном пространстве и окружающей исследуемую структуру жидкости. Разность потенциалов можно рассчитать по формуле Нернста:
где R - газовая постоянная, Т - абсолютная температура, F - число Фарадея, [К] вн. и [К] нар. - концентрация калия внутри и снаружи клетки.

Рис. 2.1.
Причина возникновения потенциала покоя общая для всех клеток. Между протоплазмой клетки и внеклеточной средой существует неравномерное распределение ионов (ионная асимметрия). Состав крови человека по солевому балансу напоминает состав океанской воды. Внеклеточная среда в центральной нервной системе также содержит много хлорида натрия. Ионный состав цитоплазмы клеток беднее. Внутри клеток в 8-10 раз меньше ионов Na + и в 50 раз меньше ионовС!". Основным катионом цитоплазмы является К + . Его концентрация внутри клетки в 30 раз выше, чем во внеклеточной среде, и приблизительно равняется внеклеточной концентрации Na Основными противоионами для К + в цитоплазме являются органические анионы, в частности анионы аспарагиновой, гистаминовой и других аминокислот. Такая асимметрия - это нарушение термодинамического равновесия. Для того чтобы восстановить его, ионы калия должны постепенно покидать клетку, а ионы натрия - стремиться в нее. Однако этого не происходит.
Первым препятствием для выравнивания разности концентраций ионов является плазматическая мембрана клетки. Она состоит из двойного слоя молекул фосфолипидов, покрытых изнутри слоем белковых молекул, а снаружи - слоем углеводов (мукополисахари- дов). Некоторая часть клеточных белков встроена непосредственно в двойной липидный слой. Это внутренние белки.
Мембранные белки всех клеток делят на пять классов: насосы, каналы, рецепторы, ферменты и структурные белки. Насосы служат для перемещения ионов и молекул против градиентов концентрации, используя для этого метаболическую энергию. Белковые каналы, или поры, обеспечивают избирательную проницаемость (диффузию) через мембрану соответствующих им по размеру ионов и молекул. Рецепторные белки, обладающие высокой специфичностью, распознают и связывают, прикрепляя к мембране, многие типы молекул, необходимых для жизнедеятельности клетки в каждый определенный момент времени. Ферменты ускоряют течение химических реакций у поверхности мембраны. Структурные белки обеспечивают соединение клеток в органы и поддержание субклеточной структуры.
Все эти белки специфичны, но не строго. В определенных условиях тот или иной белок может быть одновременно и насосом, и ферментом, и рецептором. Через каналы мембраны молекулы воды, а также соответствующие размерам пор ионы входят в клетку и выходят из нее. Проницаемость мембраны для различных катионов не одинакова и изменяется при разных функциональных состояниях ткани. В покое мембрана в 25 раз более проницаема для ионов калия, чем для ионов натрия, а при возбуждении натриевая проницаемость примерно в 20 раз превышает калиевую. В состоянии покоя равные концентрации калия в цитоплазме и натрия во внеклеточной среде должны обеспечить и равное количество положительных зарядов по обе стороны мембраны. Но поскольку проницаемость для ионов калия выше в 25 раз, то калий, выходя из клетки, делает ее поверхность все более положительно заряженной по отношению к внутренней стороне мембраны, около которой все более накапливаются слишком крупные для пор мембраны отрицательно заряженные молекулы аспарагиновой, гистаминовой и других аминокислот, «отпустивших» калий за пределы клетки, но «не дающих» ему уйти далеко благодаря своему отрицательному заряду. С внутренней стороны мембраны копятся отрицательные заряды, а с внешней - положительные. Возникает разность потенциалов. Диффузный ток ионов натрия в протоплазму из внеклеточной жидкости удерживает эту разность на уровне 60-70 мВ, не давая ей возрастать. Диффузный ток ионов натрия в покое в 25 раз слабее, чем встречный ток ионов калия. Ионы натрия, проникая внутрь клетки, снижают величину потенциала покоя, позволяя ей удерживаться на определенном уровне. Таким образом, величина потенциала покоя мышечных и нервных клеток, а также нервных волокон определяется соотношением числа положительно заряженных ионов калия, диффундирующих в единицу времени из клетки наружу, и положительно заряженных ионов натрия, диффундирующих через мембрану в противоположном направлении. Чем это соотношение выше, тем больше величина потенциала покоя, и наоборот.
Вторым препятствием, удерживающим разность потенциалов на определенном уровне, является натрий-калиевый насос (рис. 2.2). Он получил название натрий-калиевого или ионного, поскольку осуществляет активное выведение (выкачивание) из протоплазмы проникающих в нее ионов натрия и введение (нагнетание) в нее ионов калия. Источником энергии для работы ионного насоса является расщепление АТФ (аденозинтрифосфата), которое происходит под воздействием фермента аденозинтрифосфатазы, локализованного в мембране клетки и активируемого теми же ионами, т. е. калием и натрием (нагрий-калий-зависимая АТФ-аза).

Рис. 2.2.
Это крупный белок, превышающий по размеру толщину клеточной мембраны. Молекула этого белка, пронизывая мембрану насквозь, связывает с внутренней стороны преимущественно натрий и АТФ, а с наружной - калий и различные ингибиторы типа глико- зидов. При этом возникает мембранный ток. Благодаря этому току обеспечивается соответствующее направление переноса ионов. Перенос ионов происходит в три этапа. Сначала ион соединяется с молекулой переносчика, образуя комплекс ион-переносчик. Затем этот комплекс проходит через мембрану или переносит через нее заряд. В завершении - ион освобождается от переносчика на противоположной стороне мембраны. Одновременно происходит аналогичный процесс, переносящий ионы в противоположном направлении. Если насос осуществляет перенос одного иона натрия на один ион калия, то он просто поддерживает концентрационный градиент по обе стороны мембраны, но не вносит вклада в создание мембранного потенциала. Чтобы внести этот вклад, ионный насос должен переносить натрий и калий в соотношении 3:2, т. е. на 2 иона калия, поступающих в клетку, из клетки он должен выводить 3 иона натрия. Работая с максимальной нагрузкой, каждый насос способен перекачивать через мембрану около 130 ионов калия и 200 ионов натрия в секунду. Это предельная скорость. В реальных условиях работа каждого насоса регулируется в соответствии с потребностями клетки. У большинства нейронов на один квадратный микрон мембранной поверхности приходится от 100 до 200 ионных насосов. Следовательно, мембрана любой нервной клетки содержит 1 миллион ионных насосов, способных перемещать до 200 миллионов ионов натрия в секунду.
Таким образом, мембранный потенциал (потенциал покоя) создается в результате как пассивных, так и активных механизмов. Степень участия тех или иных механизмов в разных клетках неодинакова, из чего следует, что мембранный потенциал может быть неодинаковым в разных структурах. Активность насосов может зависеть от диаметра нервных волокон: чем тоньше волокно, тем отношение размера поверхности к объему цитоплазмы выше, соответственно, и активность насосов, необходимая для поддержания разницы концентраций ионов на поверхности и внутри волокна, должна быть больше. Другими словами, мембранный потенциал может зависеть от структуры нервной ткани, а значит, и от ее функционального назначения. Электрическая поляризация мембраны - главное условие, обеспечивающее возбудимость клетки. Это ее постоянная готовность к действию. Это запас потенциальной энергии клетки, который она может использовать в случае, если нервной системе понадобится ее немедленная реакция.
Между наружной поверхностью клетки и ее цитоплазмой в состоянии покоя существует разность потенциалов около 0,06-0,09 в, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называют потенциалом покоя или мембранным потенциалом. Точное измерение потенциала покоя возможно только с помощью микроэлектродов, предназначенных для внутриклеточного отведения токов, очень мощных усилителей и чувствительных регистрирующих приборов - осциллографов.
Микроэлектрод (рис. 67, 69) представляет собой тонкий стеклянный капилляр, кончик которого имеет диаметр около 1 мкм. Этот капилляр заполняют солевым раствором, погружают в него металлический электрод и соединяют с усилителем и осциллографом (рис. 68). Как только микроэлектрод прокалывает покрывающую клетку мембрану, луч осциллографа отклоняется вниз из своего исходного положения и устанавливается на новом уровне. Это свидетельствует о наличии разности потенциалов между наружной и внутренней поверхностью клеточной мембраны.

Наиболее полно происхождение потенциала покоя объясняет так называемая мембранно-ионная теория. Согласно этой теории все клетки покрыты мембраной, имеющей неодинаковую проницаемость для различных ионов. В связи с этим внутри клетки в цитоплазме в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем на поверхности. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия. Диффузия положительно заряженных ионов калия из цитоплазмы на поверхность клетки придает наружной поверхности мембраны положительный заряд.
Таким образом, поверхность клетки в покое несет на себе положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других крупных органических анионов, которые через мембрану практически не проникают (рис. 70).
Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя, то в этом участке возникает возбуждение, проявляющееся в быстром колебании мембранного потенциала и называемое потенциалом действия .
Потенциал действия можно зарегистрировать либо с помощью электродов, приложенных к внешней поверхности волокна (внеклеточное отведение), либо микроэлектрода, введенного в цитоплазму (внутриклеточное отведение).
При внеклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий период, измеряемый тысячными долями секунды, становится заряженной электроотрицательно по отношению к покоящемуся участку.


Причина возникновения потенциала действия - изменение ионной проницаемости мембраны. При раздражении проницаемость клеточной мембраны для ионов натрия повышается. Ионы натрия стремятся внутрь клетки, так как, во-первых, они заряжены положительно и их влекут внутрь электростатические силы, во-вторых, концентрация их внутри клетки невелика. В покое клеточная мембрана была малопроницаемой для ионов натрия. Раздражение изменило проницаемость мембраны, и поток положительно заряженных ионов натрия из внешней среды клетки в цитоплазму значительно превышает поток ионов калия из клетки наружу. В результате внутренняя поверхность мембраны становится заряженной положительно, а наружная вследствие потери положительно заряженных ионов натрия отрицательно. В этот момент и регистрируется пик потенциала действия.
Повышение проницаемости мембраны для ионов натрия продолжается очень короткое время. Вслед за этим в клетке возникают восстановительные процессы, приводящие к тому, что проницаемость мембраны для ионов натрия вновь понижается, а для ионов калия возрастает. Поскольку ионы калия также заряжены положительно, то, выходя из клетки, они восстанавливают исходные отношения снаружи и внутри клетки.
Накопления ионов натрия внутри клетки при многократном возбуждении ее не происходит потому, что ионы натрия эвакуируются из нее постоянно за счет действия специального биохимического механизма, называемого "натриевым насосом". Есть данные и об активном транспорте ионов калия с помощью "натрий-калиевого насоса".
Таким образом, согласно мембранно-ионной теории в происхождении биоэлектрических явлений решающее значение имеет избирательная проницаемость клеточной мембраны, обусловливающая разный ионный состав на поверхности и внутри клетки, а следовательно, и разный заряд этих поверхностей. Следует заметить, что многие положения мембранно-ионной теории все еще дискуссионны и нуждаются в дальнейшей разработке.
В 1902 году Юлиус Бернштейн выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К + , и они накапливаются в цитоплазме. Расчет величины потенциала покоя по уравнению Нернста для калиевого электрода удовлетворительно совпал с измеренным потенциалом между саркоплазмой мышцы и окружающей средой, который составлял около - 70 мВ.
Согласно теории Ю. Бернштейна, при возбуждении клетки её мембрана повреждается, и ионы К + вытекают из клетки по концентрационному градиенту до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя. Это утверждение, относящееся скорее к потенциалу действия , было опровергнуто Ходжкином и Хаксли в 1939 году.
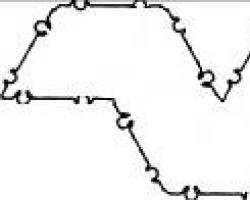
Теорию Бернштейна касательно потенциала покоя подтвердил Кеннет Стюарт Коул (Kenneth Stewart Cole), иногда его инициалы ошибочно пишут как K.C. Cole, из-за его прозвища, Кейси («Kacy»). ПП и ПД изображены на известной иллюстрации Коула и Curtis, 1939. Этот рисунок стал эмблемой Membrane Biophysics Group of the Biophysical Society (см. иллюстрацию).
Для того, чтобы на мембране поддерживалась разность потенциалов, необходимо, чтобы была определенная разность концентрации различных ионов внутри и снаружи клетки.
Концентрации ионов в клетке скелетной мышцы и во внеклеточной среде
Потенциал покоя для большинства нейронов составляет величину порядка −60 мВ - −70 мВ. У клеток невозбудимых тканей на мембране также имеется разность потенциалов, разная для клеток разных тканей и организмов.
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na + на K + в соотношении 3: 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ , обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na +) в клетке.
2. Избыток ионов калия (K +) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K + . Ионы калия K + покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 мВ.
Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
Дудель Й., Рюэгг Й., Шмидт Р. и др. Физиология человека: в 3-х томах. Пер. с англ / под ред Р. Шмидта и Г. Тевса. - 3. - М .: Мир, 2007. - Т. 1. - 323 с илл. с. - 1500 экз. - ISBN 5-03-000575-3
Wikimedia Foundation . 2010 .